
Haplotype of the Lipoprotein(a) Gene Variants rs10455872 and rs3798220 Is Associated with Parameters of Coagulation, Fibrinolysis, and Inflammation in Patients after Myocardial Infarction and Highly Elevated Lipoprotein(a) Values
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Patients’ Characteristics
2.2. Associations between LPA Genotypes, Haplotypes, and Inflammatory Parameters
2.3. Associations between LPA Genotypes, Haplotypes, and Hemostatic Parameters
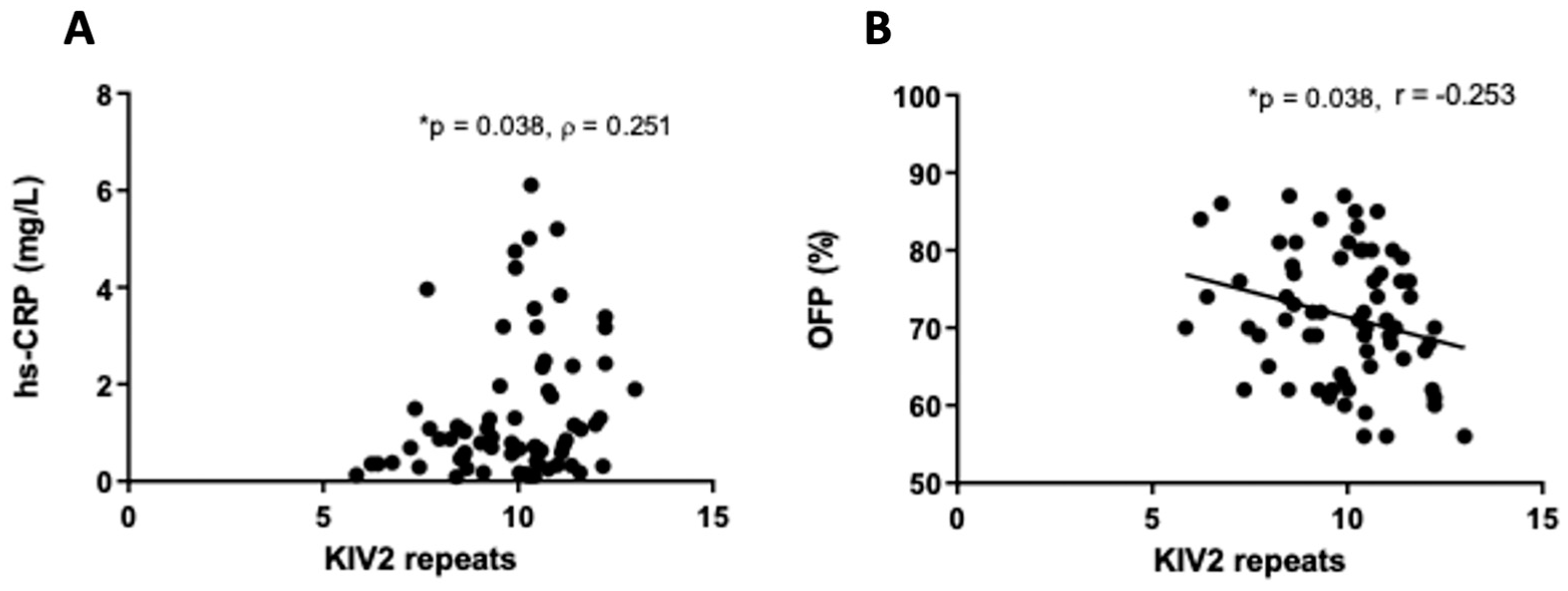
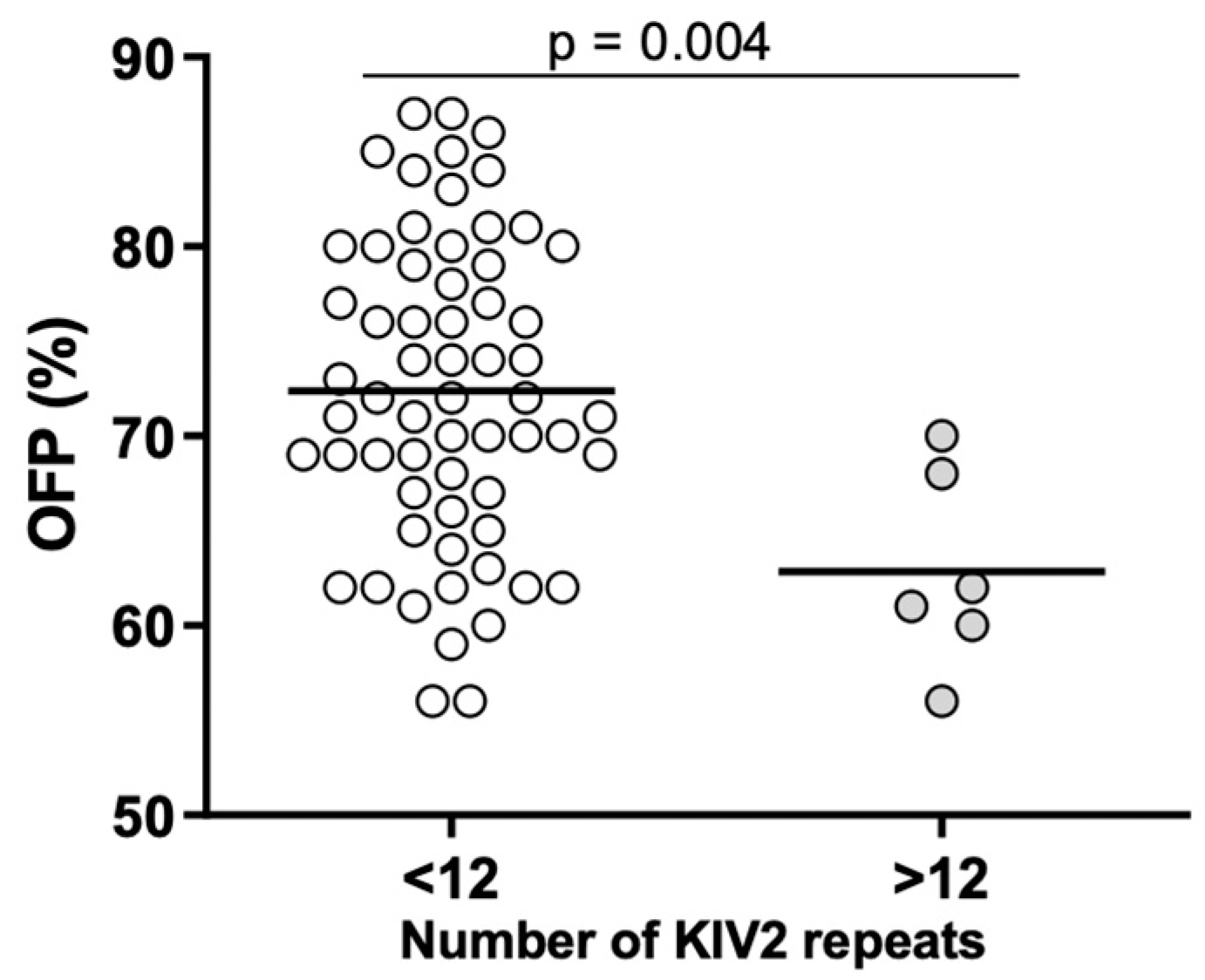
2.4. Associations between Lp(a) Levels, KIV-2 Repeats, and Inflammatory and Hemostatic Parameters
3. Discussion
4. Patients and Methods
4.1. Patients
4.2. Biochemical Analysis
4.3. Genetic Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salari, N.; Morddarvanjoghi, F.; Abdolmaleki, A.; Rasoulpoor, S.; Khaleghi, A.A.; Hezarkhani, L.A.; Shohaimi, S.; Mohammadi, M. The Global Prevalence of Myocardial Infarction: A Systematic Review and Meta—Analysis. BMC Cardiovasc. Disord. 2023, 23, 206. [Google Scholar] [CrossRef] [PubMed]
- Radšel, P. Akutni koronarni sindrom. In Interna Medicina, 5th ed.; Košnik, M., Štajer, D., Eds.; Medicinska Fakulteta Ljubljana: Ljubljana, Slovenia; Slovensko Zdravniško Društvo: Ljubljana, Slovenia, 2018; pp. 268–277. [Google Scholar]
- Kamstrup, P.R.; Benn, M.; Tybjærg-Hansen, A.; Nordestgaard, B.G. Extreme Lipoprotein(a) Levels and Risk of Myocardial Infarction in the General Population: The Copenhagen City Heart Study. Circulation 2008, 117, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Berglund, L. Lipoprotein(a) An Elusive Cardiovascular Risk Factor. Arterioscler. Thromb. Vasc. Biol. 2012, 24, 2219–2226. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Peden, J.F.; Hopewell, J.C.; Kyriakou, T.; Goel, A.; Heath, S.C.; Parish, S.; Barlera, S.; Franzosi, M.G.; Rust, S.; et al. Genetic Variants Associated with Lp(a) Lipoprotein Level and Coronary Disease. N. Engl. J. Med. 2009, 361, 2518–2528. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, X.; Feng, Q.; Kubo, M. LPA Variants Are Associated with Residual Cardiovascular Risk in Patients Receiving Statins. Circulation 2019, 138, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Bergmark, C.; Dewan, A.; Orsoni, A.; Merki, E.; Miller, E.R.; Shin, M.J.; Binder, C.J.; Hörkkö, S.; Krauss, R.M.; Chapman, M.J.; et al. A Novel Function of Lipoprotein [a] as a Preferential Carrier of Oxidized Phospholipids in Human Plasma. J. Lipid Res. 2008, 49, 2230–2239. [Google Scholar] [CrossRef] [PubMed]
- Lawn, R.M.; Schwartz, K.; Patthy, L. Convergent Evolution of Apolipoprotein(a) in Primates and Hedgehog. Proc. Natl. Acad. Sci. USA 1997, 94, 11992–11997. [Google Scholar] [CrossRef] [PubMed]
- Argraves, K.M.T.; Kozarsky, K.F.; Fallon, J.T.; Harpel, P.C.; Strickland, D.K. The Atherogenic Lipoprotein Lp(a) Is Internalized and Degraded in a Process Mediated by the VLDL Receptor. J. Clin. Investig. 1997, 100, 2170–2181. [Google Scholar] [CrossRef]
- Simantiris, S.; Antonopoulos, A.S.; Papastamos, C.; Benetos, G.; Koumallos, N.; Tsioufis, K.; Tousoulis, D. Lipoprotein(a) and Inflammation- Pathophysiological Links and Clinical Implications for Cardiovascular Disease. J. Clin. Lipidol. 2023, 17, 55–63. [Google Scholar] [CrossRef]
- Tsimikas, S.; Kiechl, S.; Willeit, J.; Mayr, M.; Miller, E.R.; Kronenberg, F.; Xu, Q.; Bergmark, C.; Weger, S.; Oberhollenzer, F.; et al. Oxidized Phospholipids Predict the Presence and Progression of Carotid and Femoral Atherosclerosis and Symptomatic Cardiovascular Disease. Five-Year Prospective Results From the Bruneck Study. J. Am. Coll. Cardiol. 2006, 47, 2219–2228. [Google Scholar] [CrossRef]
- Kiechl, S.; Willeit, J.; Mayr, M.; Viehweider, B.; Oberhollenzer, M.; Kronenberg, F.; Wiedermann, C.J.; Oberthaler, S.; Xu, Q.; Witztum, J.L.; et al. Oxidized Phospholipids, Lipoprotein(a), Lipoprotein-Associated Phospholipase A2 Activity, and 10-Year Cardiovascular Outcomes: Prospective Results from the Bruneck Study. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1788–1795. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Hammami, I.; Sherliker, P.; Valdes-Marquez, E.; Watkins, H.; Hill, M.; Yang, X.; Tsimikas, S.; Hopewell, J.C.; Consortium, P. Oxidized Phospholipids on Apolipoprotein B-100 versus Plasminogen and Risk of Coronary Heart Disease in the PROCARDIS Study. Atherosclerosis 2022, 354, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Van der Valk, F.M.; Bekkering, S.; Kroon, J.; Yeang, C.; Van den Bossche, J.; van Buul, J.D.; Ravandi, A.; Nederveen, A.J.; Verberne, H.J.; Scipione, C.; et al. Oxidized Phospholipids on Lipoprotein(a) Elicit Arterial Wall Inflammation and an Inflammatory Monocyte Response in Humans. Circulation 2016, 134, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, J.G.; Poels, K.; Stiekema, L.C.A.; Yeang, C.; Tsimikas, S.; Kroon, J.; Stroes, E.S.G.; Lutgens, E.; Seijkens, T.T.P. Short-Term Regulation of Hematopoiesis by Lipoprotein(a) Results in the Production of pro-Inflammatory Monocytes. Int. J. Cardiol. 2020, 315, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Tsironis, L.D.; Mitsios, J.V.; Milionis, H.J.; Elisaf, M.; Tselepis, A.D. Effect of Lipoprotein (a) on Platelet Activation Induced by Platelet-Activating Factor: Role of Apolipoprotein (a) and Endogenous PAF-Acetylhydrolase. Cardiovasc. Res. 2004, 63, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Coassin, S.; Kronenberg, F. Lipoprotein(a) beyond the Kringle IV Repeat Polymorphism: The Complexity of Genetic Variation in the LPA Gene. Atherosclerosis 2022, 349, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hong, C.E.; Lewis, J.P.; Zhu, Y.; Wang, X.; Chu, X.; Backman, J.; Hu, Z.; Yang, P.; Still, C.D.; et al. Effect of Two Lipoprotein (a)-Associated Genetic Variants on Plasminogen Levels and Fibrinolysis. G3 Genes Genomes Genet. 2016, 6, 3525–3532. [Google Scholar] [CrossRef]
- Scipione, C.A.; McAiney, J.T.; Simard, D.J.; Bazzi, Z.A.; Gemin, M.; Romagnuolo, R.; Macrae, F.L.; Ariëns, R.A.; Hegele, R.A.; Auld, J.; et al. Characterization of the I4399M Variant of Apolipoprotein(a): Implications for Altered Prothrombotic Properties of Lipoprotein(A); Wiley Online Library: Hoboken, NJ, USA, 2017; Volume 15, ISBN 1519931522. [Google Scholar]
- Likozar, A.R.; Blinc, A.; Podkrajšek, K.T.; Šebeštjen, M. LPA Genotypes and Haplotypes Are Associated with Lipoprotein(A) Levels but Not Arterial Wall Properties in Stable Post-Coronary Event Patients with Very High Lipoprotein(a) Levels. J. Cardiovasc. Dev. Dis. 2021, 8, 181. [Google Scholar] [CrossRef]
- Task, A.; Members, F.; Mach, F.; Baigent, C.; Catapano, A.L.; Koskina, K.C.; Casula, M.; Badimon, L.; Chapman, M.J.; De Backer, G.G.; et al. 2019 ESC/EAS Guidelines for the Management of Dyslipidaemias: Lipid Modification to Reduce Cardiovascular Risk. Atherosclerosis 2019, 290, 140–205. [Google Scholar] [CrossRef]
- Levstek, T.; Podkrajšek, N.; Likozar, A.R.; Šebeštjen, M.; Podkrajšek, K.T. The Influence of Treatment with PCSK9 Inhibitors and Variants in the CRP (Rs1800947), TNFA (Rs1800629), and IL6 (Rs1800795) Genes on the Corresponding Inflammatory Markers in Patients with Very High Lipoprotein (a) Levels. J. Cardiovasc. Dev. Dis. 2022, 9, 127. [Google Scholar] [CrossRef]
- Kamstrup, P.R.; Tybjærg-hansen, A.; Steffensen, R.; Nordestgaard, B.G. Genetically Elevated Lipoprotein (A). J. Am. Med. Assoc. 2009, 301, 2331–2339. [Google Scholar] [CrossRef]
- Arai, K.; Luke, M.M.; Koschinsky, M.L.; Miller, E.R.; Pullinger, C.R.; Witztum, J.L.; Kane, J.P.; Tsimikas, S. The I4399M Variant of Apolipoprotein(a) Is Associated with Increased Oxidized Phospholipids on Apolipoprotein B-100 Particles. Atherosclerosis 2010, 209, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Mcconnell, J.P.; Lennon, R.J.; Kornman, K.S.; Witztum, J.L.; Berger, P.B. Oxidized Phospholipids, Lp(a) Lipoprotein, and Coronary Artery Disease. N. Engl. J. Med. 2005, 353, 46–57. [Google Scholar]
- Ansari, W.M.; Humphries, S.E.; Naveed, A.K.; Khan, O.J.; Khan, D.A.; Khattak, E.H. Effect of Coronary Artery Disease Risk SNPs on Serum Cytokine Levels and Cytokine Imbalance in Premature Coronary Artery Disease. Cytokine 2019, 122, 154060. [Google Scholar] [CrossRef] [PubMed]
- Willeit, P.; Ridker, P.M.; Nestel, P.J.; Simes, J.; Tonkin, A.M.; Pedersen, T.R.; Schwartz, G.G.; Olsson, A.G.; Colhoun, H.M.; Kronenberg, F.; et al. Baseline and On-Statin Treatment Lipoprotein(a) Levels for Prediction of Cardiovascular Events: Individual Patient-Data Meta-Analysis of Statin Outcome Trials. Lancet 2018, 392, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Pirro, M.; Bianconi, V.; Paciullo, F.; Mannarino, M.R.; Bagaglia, F.; Sahebkar, A. Lipoprotein(a) and Inflammation: A Dangerous Duet Leading to Endothelial Loss of Integrity. Pharmacol. Res. 2017, 119, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Meade, T.W.; Ruddock, V.; Stirling, Y.; Chakrabarti, R.; Miller, G.J. Fibrinolytic Activity, Clotting Factors, and Long-Term Incidence of Ischaemic Heart Disease in the Northwick Park Heart Study. Lancet 1993, 342, 1076–1079. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.; Spinthakis, N.; Gue, Y.X.; Srinivasan, M.; Sullivan, K.; Wellsted, D.; Gorog, D.A. Impaired Endogenous Fibrinolysis in STEMI Patients Undergoing PPCI Is an Independent Predictor of Recurrent Cardiovascular Events: The RISK PPCI Study. Eur. Heart J. 2019, 40, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Longstaff, C. Measuring Fibrinolysis: From Research to Routine Diagnostic Assays. J. Thromb. Haemost. 2018, 16, 652–662. [Google Scholar] [CrossRef]
- He, S.; Bremme, K.; Blombäck, M. A Laboratory Method for Determination of Overall Haemostatic Potential in Plasma. I. Method Design and Preliminary Results. Thromb. Res. 1999, 96, 145–156. [Google Scholar] [CrossRef]
- Vu, M.; Bo, M.; Peternel, P.; Stegnar, M. Enhanced Thrombin Generation in Women with a History of Oral Contraception-Related Venous Thrombosis. Thromb. Res. 2013, 132, 621–626. [Google Scholar] [CrossRef]
- Ugovšek, S.; Šebeštjen, M. Lipoprotein(A)—The Crossroads of Atherosclerosis, Atherothrombosis and Inflammation. Biomolecules 2022, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Santonastaso, A.; Maggi, M.; De Jonge, H.; Scotti, C. High Resolution Structure of Human Apolipoprotein (a) Kringle IV Type 2: Beyond the Lysine Binding Site. J. Lipid Res. 2020, 61, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Hervio, L.; Durlach, V.; Girard-Globa, A.; Anglés-Cano, E. Multiple Binding with Identical Linkage: A Mechanism That Explains the Effect of Lipoprotein(a) on Fibrinolysis. Biochemistry 1995, 34, 13353–13358. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the Concentration of Low-Density Lipoprotein Cholesterol in Plasma, without Use of the Preparative Ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- El Maniani, M.; Rechchach, M.; El Mahfoudi, A.; El Moudane, M.; Sabbar, A. A Calorimetric Investigation of the Liquid Bi-Ni Alloys. J. Mater. Environ. Sci. 2016, 7, 3759–3766. [Google Scholar]
- Yahya, R.; Berk, K.; Verhoeven, A.; Bos, S.; Van Der Zee, L.; Touw, J.; Erhart, G.; Kronenberg, F.; Timman, R.; Sijbrands, E.; et al. Statin Treatment Increases Lipoprotein (a) Levels in Subjects with Low Molecular Weight Apolipoprotein (a) Phenotype. Atherosclerosis 2019, 289, 201–205. [Google Scholar] [CrossRef] [PubMed]
- German, C.A.; Liao, J.K. Understanding the Molecular Mechanisms of Statin Pleiotropic Effects. Arch. Toxicol. 2023, 97, 1529–1545. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value (n = 69) |
|---|---|
| Age (years) | 51.7 ± 8.7 |
| Body mass index (kg/m2) | 28.7 ± 8.7 |
| Systolic blood pressure (mmHg) | 128 ± 15 |
| Diastolic blood pressure (mmHg) | 77 ± 9 |
| Total cholesterol (mmol/L) | 4.24 ± 0.83 |
| HDL cholesterol (mmol/L) | 1.20 ± 0.28 |
| LDL cholesterol (mmol/L) | 2.35 ± 0.70 |
| Triglycerides (mmol/L) | 1.65 ± 0.79 |
| Lipoprotein(a) (mg/L) | 1431 (11207–1783) |
| Apolipoprotein B (g/L) | 0.81 ± 0.22 |
| Apolipoprotein A1 (g/L) | 1.34 ± 0.19 |
| KIV-2 repeats (n) | 10.2 (5.8–13.5) |
| hs-CRP (mg/L) | 0.87 (0.37–1.93) |
| TNF-α (ng/L) | 3.91 (3.36–5.36) |
| IL-6 (ng/L) | 1.69 (1.20–2.53) |
| IL-8 (ng/L) | 13.50 (10.65–18.35) |
| D-dimer (µg/L) | 270 (190–482) |
| F VIII (IU/mL) | 1.56 (1.24–1.74) |
| TAFI (%) | 100.50 (86.00–110.25) |
| PAI-1 (ng/mL) | 36.50 (22.25–56.75) |
| OCP (Abs-sum) | 23.45 (19.78–28.13) |
| OHP (Abs-sum) | 6.60 (4.73–8.65) |
| OFP (%) | 71.54 ± 8.32 |
| Parameter | Subgroup | p Value | ||
|---|---|---|---|---|
| rs10455872 | A/A (n = 40) | A/G (n = 25) | G/G (n = 5) | |
| IL-6 (ng/L) | 1.69 (1.20–2.44) | 2.07 (1.41–2.67) | 2.07 (1.41–2.67) | 0.328 |
| IL-8 (ngl/L) | 12.55 (10.43–17.30) | 14.60 (11.40–36.65) | 2.56 (0.83–5.59) | 0.383 |
| TNF-α (ng/L) | 3.91 (3.36–5.14) | 3.91 (3.21–16.12) | 4.56 (3.31–5.10) | 0.921 |
| VCAM-1 (ng/L) | 0.62(0.50–0.89) | 0.65 (0.54–0.93) | 0.52 (0.29–1.43) | 0.489 |
| rs3798220 | T/T (n = 48) | T/C (n = 21) | C/C (n = 1) | |
| IL-6 (ng/L) | 1.69 (1.17–2.67) | 1.66 (1.20–2.15) | 2.18 | 0.558 |
| IL-8 (ng/L) | 13.00 (10.50–17.48) | 16.30 (10.70–20.50) | 17.90 | 0.524 |
| TNF-α (ng/L) | 3.91 (3.13–5.57) | 4.46 (3.36–5.14) | 3.91 | 0.940 |
| VCAM-1 (ng/L) | 0.60 (0.51–0.86) | 0.78 (0.51–0.92) | 0.72 | 0.445 |
| Haplotype AT | no AT (n = 8) | one AT (n = 42) | two AT (n = 20) | |
| IL-6 (ng/L) | 1.90 (1.05–4.50) | 1.69 (1.20–2.67) | 1.45 (1.17–2.53) | 0.640 |
| IL-8 (ng/L) | 17.65 (6.90–34.80) | 14.60 (11.30–20.60) | 10.90 (10.18–15.48) | 0.281 |
| TNF-α (ng/L) | 4.56 (3.70–10.52) | 3.91 (3.36–5.14) | 3.91 (3.13–10.03) | 0.790 |
| VCAM-1 (ng/L) | 0.61 (0.40–0.97) | 0.78 (0.53–0.92) | 0.59 (0.47–0.64) | 0.087 |
| Haplotype AC | no AC (n = 48) | one AC (n = 21) | two AC (n = 1) | |
| IL-6 (ng/L) | 1.69 (1.17–2.67) | 1.66 (1.20–2.15) | 2.18 | 0.558 |
| IL-8 (ng/L) | 13.00 (10.50–17.48) | 16.30 (10.70–20.50) | 17.90 | 0.524 |
| TNF-α (ng/L) | 3.91 (3.13–5.57) | 4.46 (3.36–5.14) | 3.91 | 0.940 |
| VCAM-1 (ng/L) | 0.60 (0.51–0.86) | 0.78 (0.51–0.92) | 0.72 | 0.445 |
| Haplotype GT | no GT (n = 40) | one GT (n = 25) | two GT (n = 5) | |
| IL-6 (ng/L) | 1.69 (1.20–2.44) | 2.07 (1.41–2.67) | 2.56 (0.83–5.59) | 0.328 |
| IL-8 (ng/L) | 12.55 (10.43–17.30) | 14.60 (11.40–36.65) | 13.05 (3.30–19.95) | 0.383 |
| TNF-α (ng/L) | 3.91 (3.36–5.14) | 3.91 (3.21–16.12) | 4.56 (3.31–5.10) | 0.921 |
| VCAM-1 (ng/L) | 0.62 (0.50–0.89) | 0.65 (0.54–0.93) | 0.52 (0.29–1.43) | 0.489 |
| Parameter | Subgroup | p Value | ||
|---|---|---|---|---|
| rs10455872 | A/A (n = 40) | A/G (n = 25) | G/G (n = 4) | |
| D-dimer (µg/L) | 270.00 (190.00–550.00) | 268.00 (190.00–385.75) | 313.00 (203.75–429.00) | 0.786 |
| TAFI (%) | 102.00 (88.25–110.25) | 93.00 (86.50–110.25) | 93.50 (76.00–122.25) | 0.784 |
| OCP (Abs-sum) | 24.45 (20.03–29.60) | 23.10 (19.55–27.10) | 23.50 (16.80–24.60) | 0.715 |
| OHP (Abs-sum) | 6.50 (4.65–8.63) | 6.70 (4.70–8.60) | 6.00 (5.00–7.00) | 0.946 |
| OFP (%) | 72.00 ± 8.65 | 71.16 ± 8.18 | 68.67 ± 6.11 | 0.819 |
| rs3798220 | T/T (n = 48) | T/C (n = 20) | C/C (n = 1) | |
| D-dimer (µg/L) | 270.00 (190.00–421.00) | 295.00 (192.00–649.00) | 190.00 | 0.299 |
| TAFI (%) | 100.00 (85.00–111.00) | 100.00 (89.75–109.25) | 113.00 | 0.519 |
| OCP (Abs-sum) | 23.40 (19.60–27.80) | 24.05 (19.78–30.20) | 30.00 | 0.362 |
| OHP (Abs-sum) | 6.30 (4.70–8.30) | 7.40 (5.53–8.90) | 6.70 | 0.583 |
| OFP (%) | 71.68 ± 8.16 | 70.90 ± 8.95 | 78.00 | 0.615 |
| Haplotype AT | no AT (n = 6) | one AT (n = 43) | two AT (n = 20) | |
| D-dimer (µg/L) | 217.50 (190.00–397.00) | 279.00 (192.00–526.00) | 252.50 (190.00–484.50) | 0.460 |
| TAFI (%) | 99.00 (78.00–116.50) | 97.50 (88.25–109.75) | 102.00 (84.25–112.50) | 0.997 |
| OCP (Abs-sum) | 23.50 (19.35–31.05) | 23.40 (19.60–28.20) | 22.40 (20.20–27.73) | 0.932 |
| OHP (Abs-sum) | 6.50 (5.50–9.50) | 27.10 (4.70–8.70) | 5.60 (4.45–8.15) | 0.558 |
| OFP (%) | 70.80 ± 5.93 | 71.09 ± 8.63 | 72.70 ± 8.37 | 0.737 |
| Haplotype AC | no AC (n = 48) | one AC (n = 20) | two AC (n = 1) | |
| D-dimer (µg/L) | 270.00 (190.00–421.00) | 295.00 (192.00–649.00) | 190.00 | 0.299 |
| TAFI (%) | 100.00 (85.00–111.00) | 100.00 (89.75–109.25) | 113.00 | 0.519 |
| OCP (Abs-sum) | 23.40 (19.60–27.80) | 24.05 (19.78–30.20) | 30.00 | 0.362 |
| OHP (Abs-sum) | 6.30 (4.70–8.30) | 7.40 (5.53–8.90) | 6.70 | 0.583 |
| OFP (%) | 71.68 ± 8.16 | 70.90 ± 8.95 | 78.00 | 0.615 |
| Haplotype GT | no GT (n = 40) | one GT (n = 25) | two GT (n = 4) | |
| D-dimer (µg/L) | 270.00 (190.00–550.00) | 268.00 (190.00–385.75) | 313.00 (203.75–429.00) | 0.786 |
| TAFI (%) | 102.00 (88.25–110.25) | 93.00 (86.50–110.25) | 93.50(76.00–122.25) | 0.784 |
| OCP (Abs-sum) | 24.45 (20.03–29.60) | 23.10 (19.55–27.10) | 23.50 (16.80–27.40) | 0.715 |
| OHP (Abs-sum) | 6.50 (4.65–8.63) | 6.70 (4.70–8.60) | 6.00 (5.00–7.00) | 0.946 |
| OFP (%) | 72.00 ± 8.65 | 71.16 ± 8.18 | 68.67 ± 6.11 | 0.819 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ugovšek, S.; Rehberger Likozar, A.; Levstek, T.; Trebušak Podkrajšek, K.; Zupan, J.; Šebeštjen, M. Haplotype of the Lipoprotein(a) Gene Variants rs10455872 and rs3798220 Is Associated with Parameters of Coagulation, Fibrinolysis, and Inflammation in Patients after Myocardial Infarction and Highly Elevated Lipoprotein(a) Values. Int. J. Mol. Sci. 2024, 25, 736. https://doi.org/10.3390/ijms25020736
Ugovšek S, Rehberger Likozar A, Levstek T, Trebušak Podkrajšek K, Zupan J, Šebeštjen M. Haplotype of the Lipoprotein(a) Gene Variants rs10455872 and rs3798220 Is Associated with Parameters of Coagulation, Fibrinolysis, and Inflammation in Patients after Myocardial Infarction and Highly Elevated Lipoprotein(a) Values. International Journal of Molecular Sciences. 2024; 25(2):736. https://doi.org/10.3390/ijms25020736
Chicago/Turabian StyleUgovšek, Sabina, Andreja Rehberger Likozar, Tina Levstek, Katarina Trebušak Podkrajšek, Janja Zupan, and Miran Šebeštjen. 2024. "Haplotype of the Lipoprotein(a) Gene Variants rs10455872 and rs3798220 Is Associated with Parameters of Coagulation, Fibrinolysis, and Inflammation in Patients after Myocardial Infarction and Highly Elevated Lipoprotein(a) Values" International Journal of Molecular Sciences 25, no. 2: 736. https://doi.org/10.3390/ijms25020736