
Grape ASR-Silencing Sways Nuclear Proteome, Histone Marks and Interplay of Intrinsically Disordered Proteins

 and
and
Abstract
:1. Introduction
2. Results
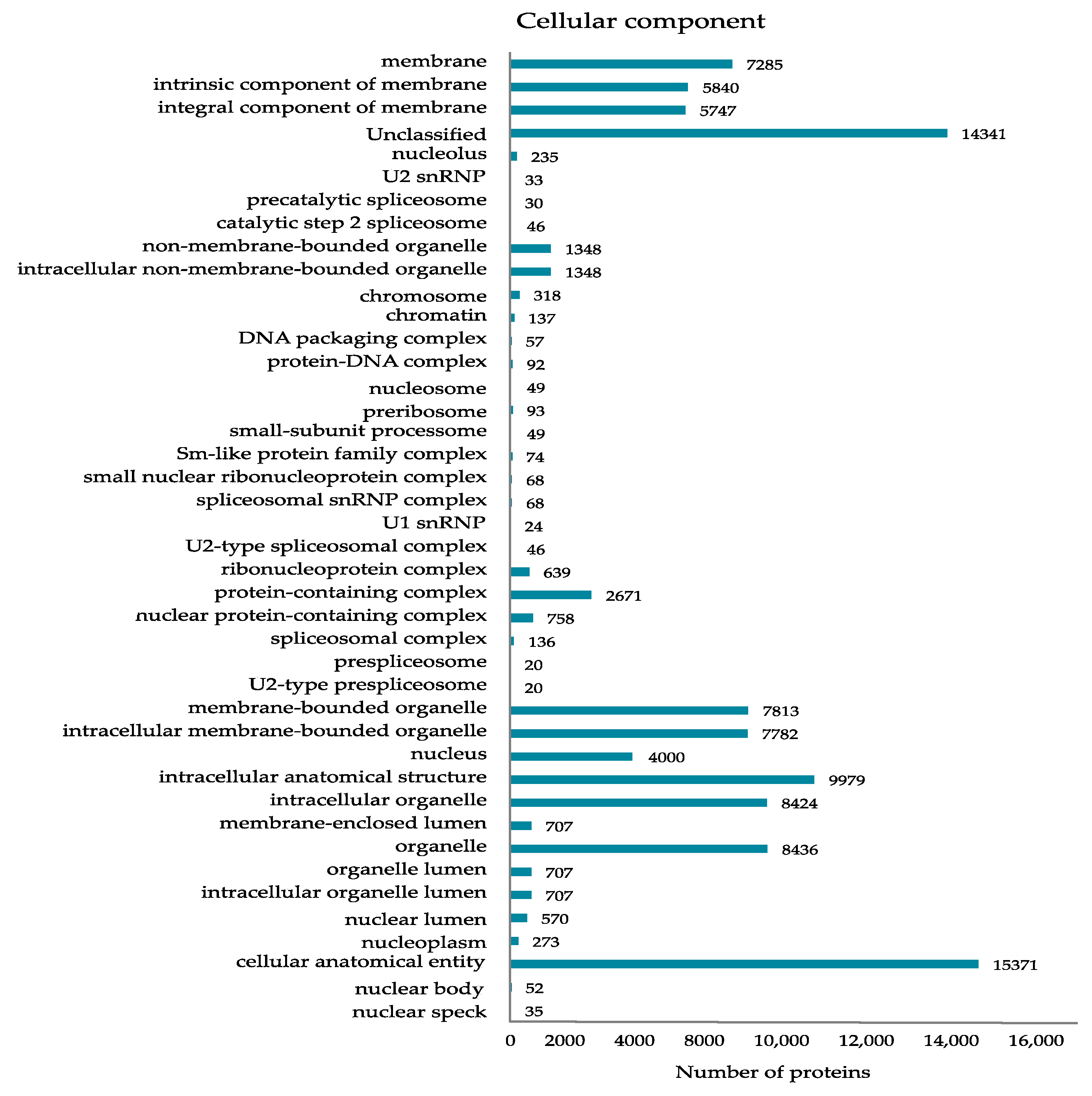
2.1. Nuclear Proteome of Grape Wild-Type and VvMSA-RNAi-Silenced Cells
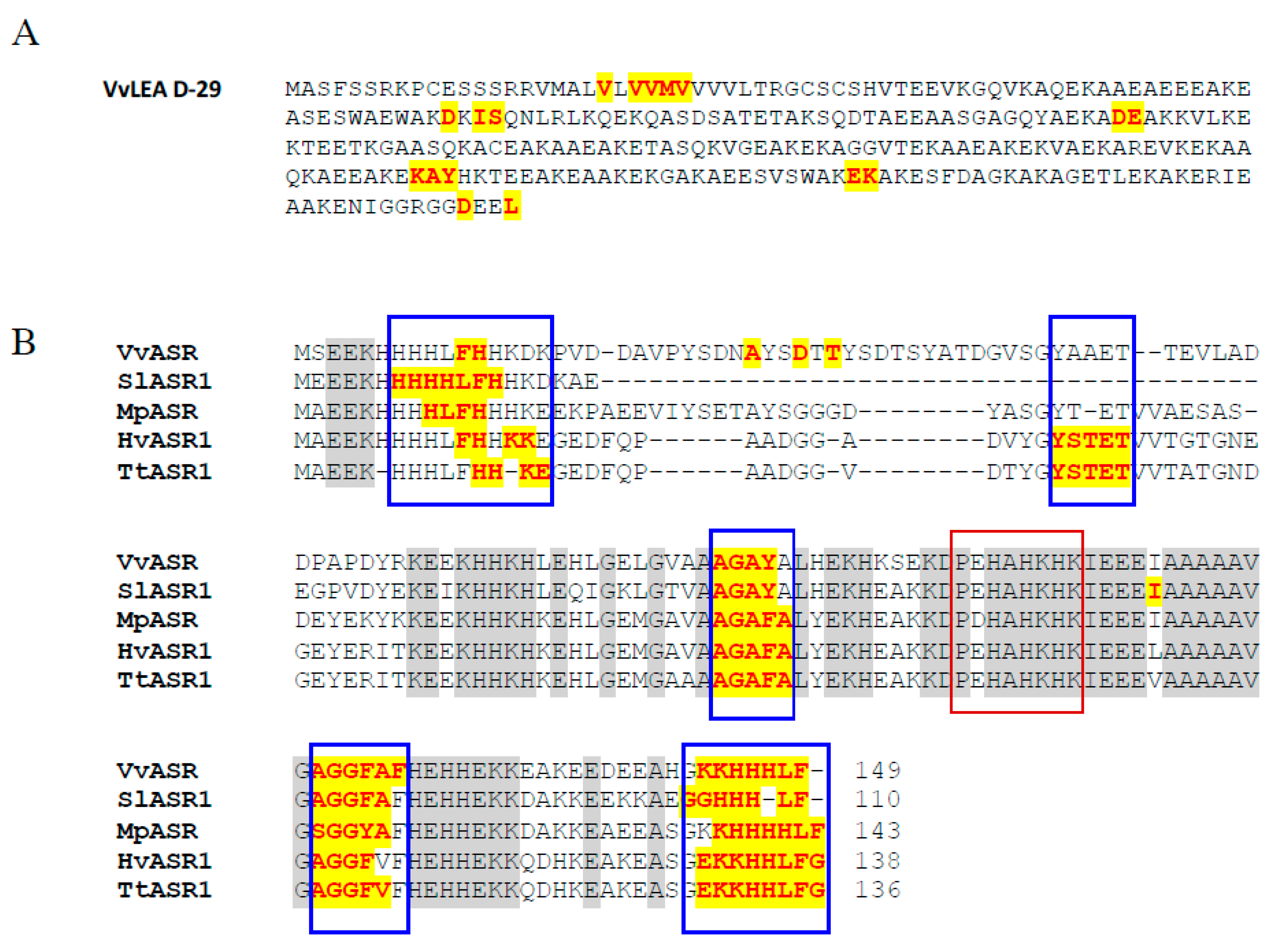
2.2. Late Embryogenesis Abundant Protein VvLEA-D29
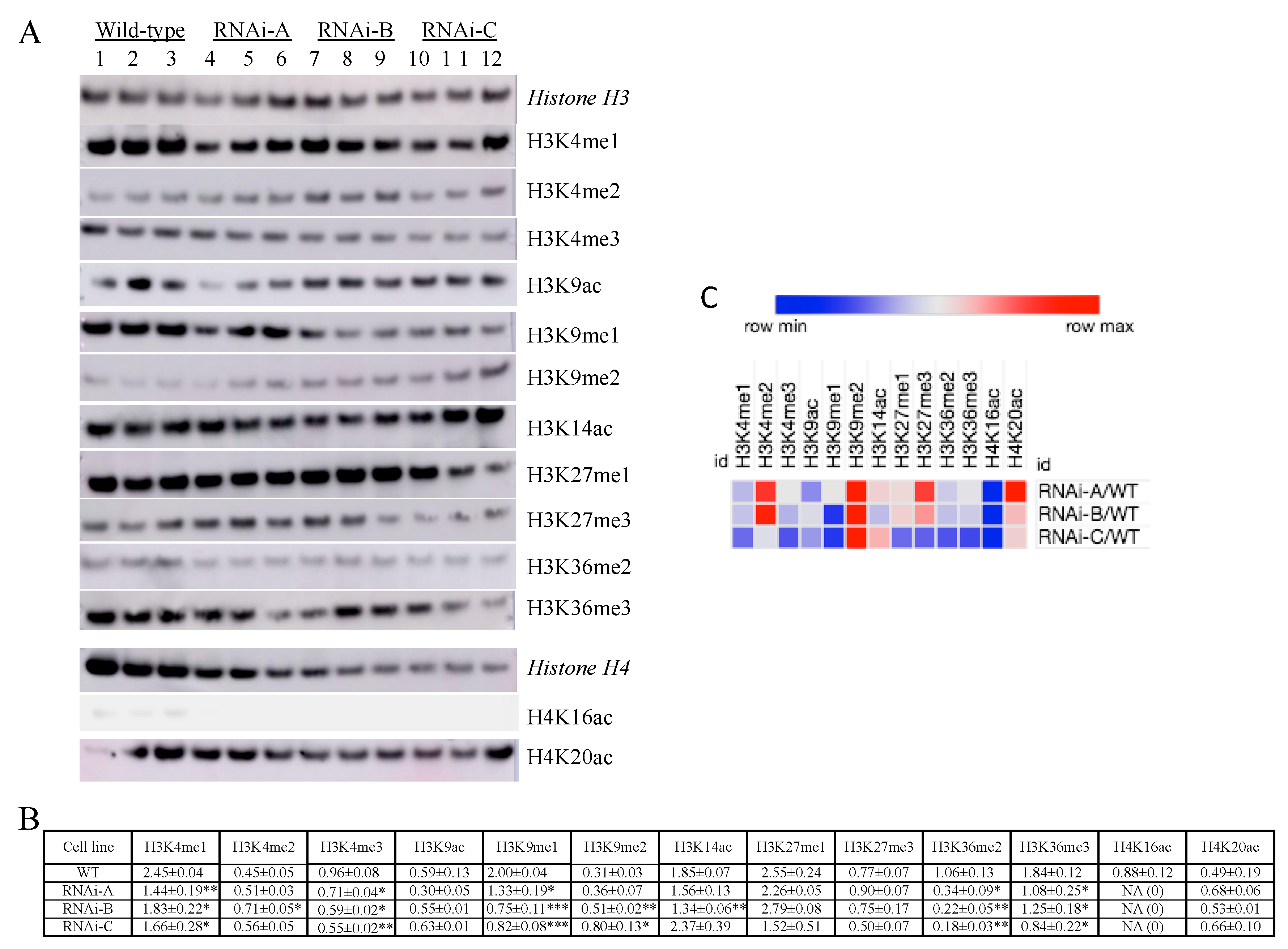
2.3. Impact of VvMSA Repression on H3 and H4 Histone Post-Translational Modifications (HPTMs)
3. Discussion
4. Materials and Methods
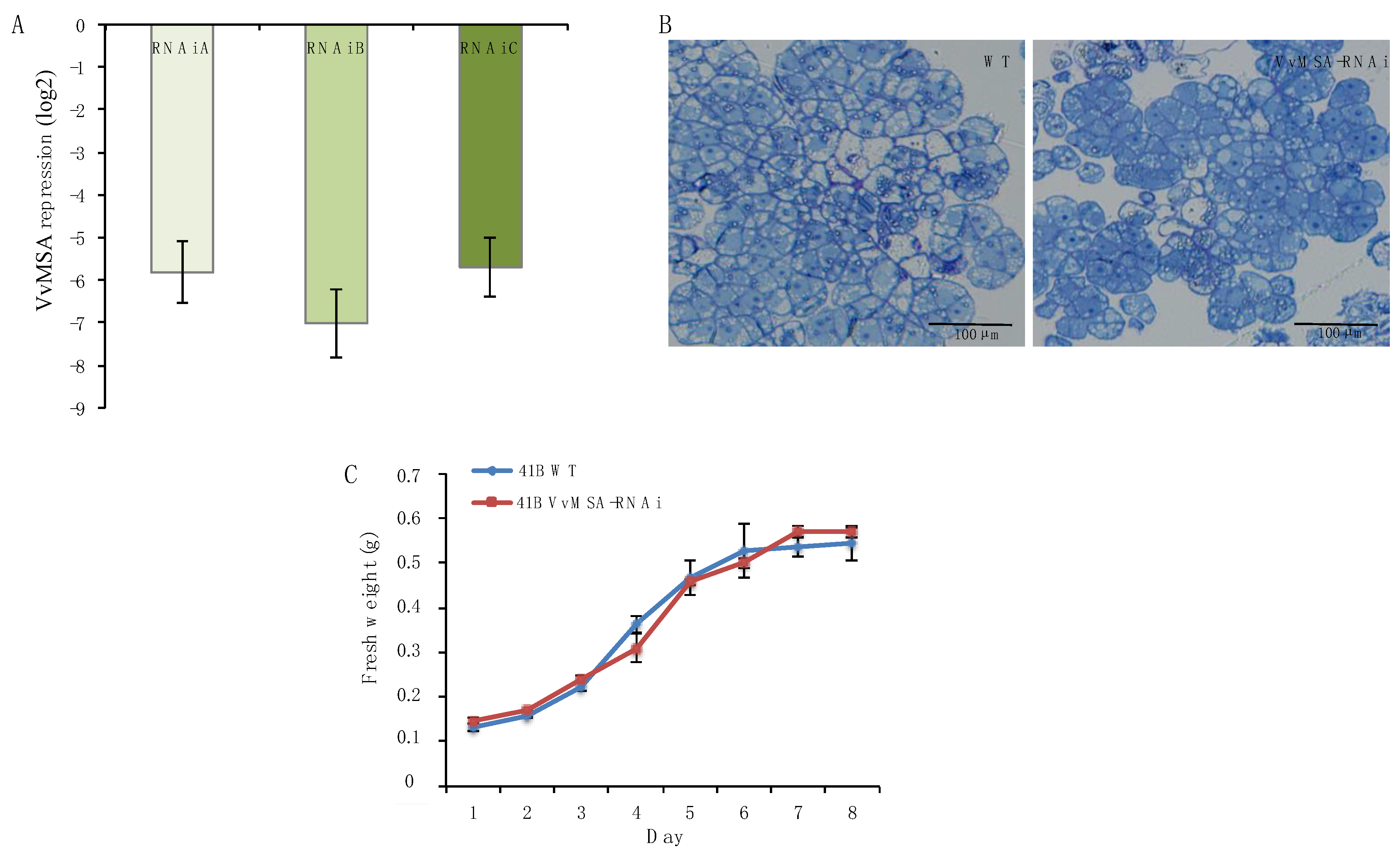
4.1. Cell Culture and Transformation Conditions
4.2. Real-Time qPCR Analysis
4.3. Isolation of Nuclei and Extraction of Nuclear Proteins
4.4. Experimental Design, Relative Quantification of Protein Abundance and Statistics
4.5. Protein Digestion, iTRAQ 8-Plex Labeling, and Peptide Fractionation
4.6. Liquid Chromatography and Tandem Mass Spectrometry
4.7. Peak List Generation and Database Search
4.8. Immunoblotting Analysis
4.9. In Silico Protein Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iusem, N.D.; Bartholomew, D.M.; Hitz, W.D.; Scolnik, P.A. Tomato (Lycopersicon esculentum) transcript induced by water deficit and ripening. Plant Physiol. 1993, 102, 1353–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, A.; Salamini, F.; Gebhardt, C. Expression patterns and promoter activity of the cold-regulated gene ci21A of potato. Plant Physiol. 1997, 113, 335–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.S.; Liau, Y.E.; Huang, J.C.; Wu, T.D.; Su, C.C.; Lin, C.H. Characterization of a desiccation-related protein in lily pollen during development and stress. Plant Cell Physiol. 1998, 39, 1307–1314. [Google Scholar] [CrossRef] [PubMed]
- de Vienne, D.; Leonardi, A.; Damerval, C.; Zivy, M. Genetics of proteome variation for QTL characterization: Application to drought-stress responses in maize. J. Exp. Bot. 1999, 50, 303–309. [Google Scholar] [CrossRef]
- Vaidyanathan, R.; Kuruvilla, S.; Thomas, G. Characterization and expression pattern of an abscisic acid and osmotic stress responsive gene from rice. Plant Sci. 1999, 140, 21–30. [Google Scholar] [CrossRef]
- Jeanneau, M.; Gerentes, D.; Foueillassar, X.; Zivy, M.; Vidal, J.; Toppan, A.; Perez, P. Improvement of drought tolerance in maize: Towards the functional validation of the Zm-Asr1 gene and increase of water use efficiency by over-expressing C4–PEPC. Biochimie 2002, 84, 1127–1135. [Google Scholar] [CrossRef]
- Liu, H.Y.; Dai, J.R.; Feng, D.R.; Liu, B.; Wang, H.B.; Wang, J.F. Characterization of a novel plantain Asr gene, MpAsr, that is regulated in response to infection of Fusarium oxysporum f. sp. cubense and abiotic stresses. J. Integr. Plant Biol. 2010, 52, 315–323. [Google Scholar] [CrossRef]
- Arenhart, R.A.; Lima, J.C.; Pedron, M.; Carvalho, F.E.; Silveira, J.A.; Rosa, S.B.; Caverzan, A.; Andrade, C.M.B.; Schünemann, M.; Margis, R.; et al. Involvement of ASR genes in aluminium tolerance mechanisms in rice. Plant Cell Environ. 2013, 36, 52–67. [Google Scholar] [CrossRef]
- Medici, A.; Laloi, M.; Atanassova, R. Profiling of sugar transporter genes in grapevine coping with water deficit. FEBS Lett. 2014, 588, 3989–3997. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Zhong, Y.; Li, Y.; Zheng, D.; Cheng, Z.M. Genome-wide identification and expression analysis of the apple ASR gene family in response to Alternaria alternata f. sp. mali. Genome 2016, 59, 866–878. [Google Scholar] [CrossRef] [Green Version]
- Cakir, B.; Agasse, A.; Gaillard, C.; Saumonneau, A.; Delrot, S.; Atanassova, R. A grape ASR protein involved in sugar and ABA signaling. Plant Cell 2003, 15, 2165–2180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalifa, Y.; Gilad, A.; Konrad, Z.; Zaccai, M.; Scolnik, P.A.; Bar-Zvi, D. The water-and salt-stress-regulated Asr1 (abscisic acid stress ripening) gene encodes a zinc-dependent DNA-binding protein. Biochem. J. 2004, 381, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.J.; Hsu, C.M.; Jauh, G.Y.; Wang, C.S. A lily pollen ASR protein localizes to both cytoplasm and nuclei requiring a nuclear localization signal. Physiol. Plant. 2005, 123, 314–320. [Google Scholar] [CrossRef]
- Ricardi, M.M.; Guaimas, F.F.; González, R.M.; Burrieza, H.P.; López-Fernández, M.P.; Jares-Erijman, E.A.; Estévez, J.M.; Iusem, N.D. Nuclear import and dimerization of tomato ASR1, a water stress-inducible protein exclusive to plants. PLoS ONE 2012, 7, e41008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenhart, R.A.; Schunemann, M.; Bucker Neto, L.; Margis, R.; Wang, Z.Y.; Margis-Pinheiro, M. Rice ASR1 and ASR5 are complementary transcription factors regulating aluminium responsive genes. Plant Cell Environ. 2016, 39, 645–651. [Google Scholar] [CrossRef]
- Hsu, Y.F.; Yu, S.C.; Yang, C.Y.; Wang, C.S. Lily ASR protein-conferred cold and freezing resistance in Arabidopsis. Plant Physiol. Biochem. 2011, 49, 937–945. [Google Scholar] [CrossRef]
- Konrad, Z.; Bar-Zvi, D. Synergism between the chaperone-like activity of the stress regulated ASR1 protein and the osmolyte glycine-betaine. Planta 2008, 227, 1213–1219. [Google Scholar] [CrossRef]
- Frankel, N.; Nunes-Nesi, A.; Balbo, I.; Mazuch, J.; Centeno, D.; Iusem, N.D.; Fernie, A.R.; Carrari, F. ci21A/Asr1expression influences glucose accumulation in potato tubers. Plant Mol. Biol. 2007, 63, 719–730. [Google Scholar] [CrossRef]
- Dominguez, P.G.; Frankel, N.; Mazuch, J.; Balbo, I.; Iusem, N.; Fernie, A.R.; Carrari, F. ASR1 mediates glucose-hormone cross talk by affecting sugar trafficking in tobacco plants. Plant Physiol. 2013, 161, 1486–1500. [Google Scholar] [CrossRef] [Green Version]
- Saumonneau, A.; Laloi, M.; Lallemand, M.; Rabot, A.; Atanassova, R. Dissection of the transcriptional regulation of grape ASR and response to glucose and abscisic acid. J. Exp. Bot. 2012, 63, 1495–1510. [Google Scholar] [CrossRef] [Green Version]
- Fàbregas, N.; Fernie, A.R. The interface of central metabolism with hormone signaling in plants. Curr. Biol. 2021, 31, R1535–R1548. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.; Olvera-Carrillo, Y.; Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The enigmatic LEA proteins and other hydrophilins. Plant Physiol. 2008, 148, 6–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldgur, Y.; Rom, S.; Ghirlando, R.; Shkolnik, D.; Shadrin, N.; Konrad, Z.; Bar-Zvi, D. Desiccation and zinc binding induce transition of tomato abscisic acid stress ripening 1, a water stress- and salt stress-regulated plant-specific protein, from unfolded to folded state. Plant Physiol. 2007, 143, 617–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.R.; Liu, B.; Feng, D.R.; Liu, H.Y.; He, Y.M.; Qi, K.B.; Wang, H.B.; Wang, J.F. MpAsr encodes an intrinsically unstructured protein and enhances osmotic tolerance in transgenic Arabidopsis. Plant Cell Rep. 2011, 30, 1219–1230. [Google Scholar] [CrossRef]
- Hamdi, K.; Salladini, E.; O’Brien, D.P.; Brier, S.; Chenal, A.; Yacoubi, I.; Longhi, S. Structural disorder and induced folding within two cereal, ABA stress and ripening (ASR) proteins. Sci. Rep. 2017, 7, 15544. [Google Scholar] [CrossRef]
- Wright, P.E.; Dyson, H.J. Intrinsically unstructured proteins: Re-assessing the protein structure-function paradigm. J. Mol. Biol. 1999, 293, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Uversky, V.N.; Gillespie, J.R.; Fink, A.L. Why are “natively unfolded” proteins unstructured under physiologic conditions? Proteins 2000, 41, 415–427. [Google Scholar] [CrossRef]
- Romero, P.; Obradovic, Z.; Li, X.; Garner, E.C.; Brown, C.J.; Dunker, A.K. Sequence complexity of disordered protein. Proteins 2001, 42, 38–48. [Google Scholar] [CrossRef]
- Uversky, V.N. What does it mean to be natively unfolded? Eur. J. Biochem. 2002, 269, 2–12. [Google Scholar] [CrossRef]
- Tompa, P. Intrinsically unstructured proteins. Trends Biochem. Sci. 2002, 27, 527–533. [Google Scholar] [CrossRef]
- Maskin, L.; Frankel, N.; Gudesblat, G.; Demergasso, M.J.; Pietrasanta, L.I.; Iusem, N.D. Dimerization and DNAbinding of ASR1, a small hydrophilic protein abundant in plant tissues suffering from water loss. Biochem. Biophys. Res. Commun. 2007, 352, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Saumonneau, A.; Agasse, A.; Bidoyen, M.T.; Lallemand, M.; Cantereau, A.; Medici, A.; Laloi, M.; Atanassova, R. Interaction of grape ASR proteins with a DREB transcription factor in the nucleus. FEBS Lett. 2008, 582, 3281–3287. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.S.; Seo, Y.W. BdERF96 interacts with BdASR1 to specifically respond to drought and oxidative stress in Brachypodium distachyon. J. Plant Biochem. Biotechnol. 2021, 30, 287–296. [Google Scholar] [CrossRef]
- Ricardi, M.M.; González, R.M.; Zhong, S.; Domínguez, P.G.; Duffy, T.; Turjanski, P.G.; Salter, J.D.S.; Alleva, K.; Carrari, F.; Gionvannoni, J.J.; et al. Genome-wide data (ChIP-seq) enabled identification of cell wall-related and aquaporin genes as targets of tomato ASR1, a drought stress-responsive transcription factor. BMC Plant Biol. 2014, 14, 29. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Hu, W.; Wang, Y.; Feng, R.; Zhang, Y.; Liu, J.; Jia, C.; Miao, H.; Zhang, J.; Xu, B.; et al. The MaASR gene as a crucial component in multiple drought stress response pathways in Arabidopsis. Funct. Integr. Genom. 2015, 15, 247–260. [Google Scholar] [CrossRef]
- Neto, L.B.; Arenhart, R.A.; de Oliveira, L.F.; de Lima, J.C.; Bodanese-Zanettini, M.H.; Margis, R.; Margis-Pinheiro, M. ASR5 is involved in the regulation of miRNA expression in rice. Plant Cell Rep. 2015, 34, 1899–1907. [Google Scholar] [CrossRef]
- Arenhart, R.A.; Bai, Y.; de Oliveira, L.F.; Neto, L.B.; Schunemann, M.; Maraschin Fdos, S.; Mariath, J.; Silverio, A.; Sachetto-Martins, G.; Margis, R.; et al. New insights into aluminum tolerance in rice: The ASR5 protein binds the STAR1 promoter and other aluminum-responsive genes. Mol. Plant 2014, 7, 709–721. [Google Scholar] [CrossRef] [Green Version]
- Gilad, A.; Amitai-Zeigerson, H.; Bar-Zvi, D.; Scolnik, P.A. Asr1, a tomato water-stress regulated gene: Genomic organization, developmental regulation and DNA-binding activity. Acta Hortic. 1997, 447, 441–453. [Google Scholar] [CrossRef]
- Marín, M.; Ott, T. Intrinsic disorder in plant proteins and phytopathogenic bacterial effectors. Chem. Rev. 2014, 114, 6912–6932. [Google Scholar] [CrossRef]
- Candat, A.; Paszkiewicz, G.; Neveu, M.; Gautier, R.; Logan, D.C.; Avelange-Macherel, M.H.; Macherel, D. The ubiquitous distribution of late embryogenesis abundant proteins across cell compartments in Arabidopsis offers tailored protection against abiotic stress. Plant Cell 2014, 26, 3148–3166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rom, S.; Gilad, A.; Kalifa, Y.; Konrad, Z.; Karpasas, M.M.; Goldgur, Y.; Bar-Zvi, D. Mapping the DNA- and zinc-binding domains of ASR1 (abscisic acid stress ripening), an abiotic-stress regulated plant specific protein. Biochimie 2006, 88, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Leung, J.; Gaudin, V. Who Rules the Cell? An Epi-Tale of Histone, DNA, RNA, and the Metabolic Deep State. Front. Plant Sci. 2020, 11, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto, T.; Suzuki, Y.; Yamaguchi, I. Effects of partial suppression of ribosomal protein S6 on organ formation in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2002, 66, 2437–2443. [Google Scholar] [CrossRef] [Green Version]
- Deprost, D.; Yao, L.; Sormani, R.; Moreau, M.; Leterreux, G.; Nicolaï, M.; Bedu, M.; Robaglia, C.; Meyer, C. The Arabidopsis TOR kinase links plant growth, yield, stress resistance and mRNA translation. EMBO Rep. 2007, 8, 864–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinbothe, C.; Pollmann, S.; Reinbothe, S. Singlet oxygen signaling links photosynthesis to translation and plant growth. Trends Plant Sci. 2010, 15, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Mahfouz, M.M.; Kim, S.; Delauney, A.J.; Verma, D.P.S. Arabidopsis TARGET OF RAPAMYCIN interacts with RAPTOR, which regulates the activity of S6 kinase in response to osmotic stress signals. Plant Cell 2006, 18, 477–490. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Zhang, C.; Dittman, J.D.; Whitham, S.A. Differential requirement of ribosomal protein S6 by plant RNA viruses with different translation initiation strategies. Virology 2009, 390, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lan, P.; Gao, H.; Zheng, L.; Li, W.; Schmidt, W. Expression changes of ribosomal proteins in phosphate- and iron-deficient Arabidopsis roots predict stress-specific alterations in ribosome composition. BMC Genom. 2013, 14, 783. [Google Scholar] [CrossRef] [Green Version]
- Parrilla, J.; Gaillard, C.; Verbeke, J.; Maucourt, M.; Aleksandrov, R.A.; Thibault, F.; Fleurat-Lessard, P.; Gibon, Y.; Rolin, D.; Atanassova, R. Comparative metabolomics and glycolysis enzyme profiling of embryogenic and nonembryogenic grape cells. FEBS Open Bio 2018, 8, 784–798. [Google Scholar] [CrossRef] [Green Version]
- Dirk, L.M.A.; Abdel, C.G.; Ahmad, I.; Neta, I.C.S.; Pereira, C.C.; Pereira, F.E.C.B.; Unêda-Trevisoli, S.H.; Pinheiro, D.G.; Downie, A.B. Late embryogenesis abundant protein-client protein interactions. Plants 2020, 9, 814. [Google Scholar] [CrossRef]
- Ibrahime, M.; Kibar, U.; Kazan, K.; Özmen, C.Y.; Mutaf, F.; Aşçi, S.D.; Çakir Aydemir, B.; Ergül, A. Genome-wide identification of the LEA protein gene family in grapevine (Vitis vinifera L.). Tree Genet. Genomes 2019, 15, 55. [Google Scholar] [CrossRef]
- Ferron, F.; Longhi, S.; Canard, B.; Karlin, D. A practical overview of protein disorder prediction methods. Proteins-Struct. Funct. Bioinform. 2006, 65, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Longhi, S.; Lieutaud, P.; Canard, B. Conformational disorder. Meth. Mol. Biol. 2010, 609, 307–325. [Google Scholar] [CrossRef]
- Lieutaud, P.; Ferron, F.; Longhi, S. Predicting Conformational Disorder. Meth. Mol. Biol. 2016, 1415, 265–299. [Google Scholar] [CrossRef]
- Wise, M.J.; Tunnacliffe, A. POPP the question: What do LEA proteins do? Trends Plant Sci. 2004, 9, 13–17. [Google Scholar] [CrossRef]
- Chen, Y.; Li, C.; Zhang, B.; Yi, J.; Yang, Y.; Kong, C.; Lei, C.; Gong, M. The role of the Late Embryogenesis-Abundant (LEA) protein family in development and the abiotic stress response: A comprehensive expression analysis of potato (Solanum tuberosum). Genes 2019, 10, 148. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Liu, N.; Virlouvet, L.; Riethoven, J.J.; Fromm, M.; Avramova, Z. Four distinct types of dehydration stress memory genes in Arabidopsis thaliana. BMC Plant Biol. 2013, 13, 229. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Sridhar, V.V.; Zhu, J.; Kapoor, A.; Zhu, J.K. Distinctive core histone posttranslational modification patterns in Arabidopsis thaliana. PLoS ONE 2007, 2, e1210. [Google Scholar] [CrossRef]
- Lauria, M.; Rossi, V. Epigenetic control of gene regulation in plants. Biochim. Biophys. Acta 2011, 1809, 369–378. [Google Scholar] [CrossRef]
- Roudier, F.; Ahmed, I.; Bérard, C.; Sarazin, A.; Mary-Huard, T.; Cortijo, S.; Bouyer, D.; Caillieux, E.; Duvernois-Berthet, E.; Al-Shikhley, L.; et al. Integrative epigenomic mapping defines four main chromatin states in Arabidopsis. EMBO J. 2011, 30, 1928–1938. [Google Scholar] [CrossRef] [Green Version]
- Sani, E.; Herzyk, P.; Perrella, G.; Colot, V.; Amtmann, A. Hyperosmotic priming of Arabidopsis seedlings establishes a long-term somatic memory accompanied by specific changes of the epigenome. Genome Biol. 2013, 14, R59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asensi-Fabado, M.A.; Amtmann, A.; Perrella, G. Plant responses to abiotic stress: The chromatin context of transcriptional regulation. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 106–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lämke, J.; Brzezinka, K.; Altmann, S.; Bäurle, I. A hit-and-run heat shock factor governs sustained histone methylation and transcriptional stress memory. EMBO J. 2016, 35, 162–175. [Google Scholar] [CrossRef] [Green Version]
- González, R.M.; Ricardi, M.M.; Iusem, N.D. Epigenetic marks in an adaptive water stress-responsive gene in tomato roots under normal and drought conditions. Epigenetics 2013, 8, 864–872. [Google Scholar] [CrossRef] [Green Version]
- Ito, H.; Gaubert, H.; Bucher, E.; Mirouze, M.; Vaillant, I.; Paszkowski, J. An siRNA pathway prevents transgenerational retrotransposition in plants subjected to stress. Nature 2011, 472, 115–119. [Google Scholar] [CrossRef]
- McCue, A.D.; Nuthikattu, S.; Reeder, S.H.; Slotkin, R.K. Gene expression and stress response mediated by the epigenetic regulation of a transposable element small RNA. PLoS Genet. 2012, 8, e1002474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shia, W.J.; Pattenden, S.G.; Workman, J.L. Histone H4 lysine 16 acetylation breaks the genome’s silence. Genome Biol. 2006, 7, 217. [Google Scholar] [CrossRef] [Green Version]
- Vaquero, A.; Sternglanz, R.; Reinberg, D. NAD+-dependent deacetylation of H4 lysine 16 by class III HDACs. Oncogene 2007, 26, 5505–5520. [Google Scholar] [CrossRef] [Green Version]
- Shogren-Knaak, M.; Ishii, H.; Sun, J.M.; Pazin, M.J.; Davie, J.R.; Peterson, C.L. Histone H4-K16 acetylation controls chromatin structure and protein interactions. Science 2006, 311, 844–847. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Chen, X.; Sanders, D.; Qian, S.; Zhong, X. High-resolution mapping of H4K16 and H3K23 acetylation reveals conserved and unique distribution patterns in Arabidopsis and rice. Epigenetics 2015, 10, 1044–1053. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Zhan, Z.; Jiang, D. Histone modifications and their regulatory roles in plant development and environmental memory. J. Genet. Genom. 2019, 46, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Shaik, R.; Ramakrishna, W. Bioinformatic analysis of epigenetic and microRNA mediated regulation of drought responsive genes in rice. PLoS ONE. 2012, 7, e49331. [Google Scholar] [CrossRef]
- Coutos-Thévenot, P.; Poinssot, B.; Bonomelli, A.; Yean, H.; Breda, C.; Buffard, D.; Esnault, R.; Hain, R.; Boulay, M. In vitro tolerance to Botrytis cinerea of grapevine 41B rootstock in transgenic plants expressing the stilbene synthase Vst1 gene under the control of a pathogen-inducible PR 10 promoter. J. Exp. Bot. 2001, 52, 901–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzozie, A.; Nanni, P.; Staiano, T.; Grossmann, J.; Barkow-Oesterreicher, S.; Shay, J.W.; Tiwari, A.; Buffoli, F.; Laczko, E.; Marra, G. Sorbitol dehydrogenase overexpression and other aspects of dysregulated protein expression in human precancerous colorectal neoplasms: A quantitative proteomics study. Mol. Cell. Proteom. 2014, 13, 1198–1218. [Google Scholar] [CrossRef] [Green Version]
- Barkow-Oesterreicher, S.; Türker, C.; Panse, C. FCC - An automated rule-based processing tool for life science data. Source Code Biol. Med. 2013, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Käll, L.; Storey, J.D.; MacCoss, M.J.; Noble, W.S. Assigning significance to peptides identified by tandem mass spectrometry using decoy databases. J. Proteome Res. 2008, 7, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Disfani, F.M.; Hsu, W.L.; Mizianty, M.J.; Oldfield, C.J.; Xue, B.; Dunker, A.K.; Uversky, V.N.; Kurgan, L. MoRFpred, a computational tool for sequence-based prediction and characterization of short disorder-to-order transitioning binding regions in proteins. Bioinformatics 2012, 28, i75–i83. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProtKB Accession | NCBI Accession | STRING Accession | Protein Function | VvMSA-RNAi vs. WT |
|---|---|---|---|---|
| Cell division/Differentiation (n = 11) | ||||
| F6GVS4 (F6GVS4 _VITVI) | CBI16879.3 | VIT_14s0083g00450.t01 | Protein FLX-like 2 | 1.78 |
| D7TD96 (D7TD96_VITVI) | CBI28469.3 | VIT_12s0057g01280.t01 | G-strand specific single-stranded telomere-binding protein 2 | 1.24 |
| F6HCE0 (F6HCE0_VITVI) | CBI25499.3 | VIT_13s0067g00550.t01 | Clathrin light chain 2-like | 1.24 |
| D7T0X4 (D7T0X4_VITVI) | CBI24194.3 | VIT_19s0085g00200.t01 | Stigma/style cell cycle inhibitor 1 | 1.28 |
| F6H9W8 (F6H9W8_VITVI) | CBI22388.3 | VIT_19s0085g00220.t01 | Cell division cycle 5-like protein | 1.22 |
| D7SJS1 (D7SJS1_VITVI) | CBI15897.3 | VIT_06s0004g07170.t01 | Structural maintenance of chromosomes domain-containing protein | 1.21 |
| F6GZE1 (F6GZE1_VITVI) | CBI18813.3 | VIT_00s0920g00020.t01 | Sister chromatid cohesion protein PDS5 homolog B | 1.28 |
| D7U294 (D7U294_VITVI) | CBI36860.3 | VIT_07s0005g00470.t01 | Peter Pan-like protein | 0.38 |
| D7SJV2 (D7SJV2_VITVI) | CBI15928.3 | VIT_06s0004g06870.t01 | Proliferation-associated protein 2G4-like | 0.33 |
| D7T8K7 (D7T8K7_VITVI) | CBI26828.3 | VIT_01s0011g05790.t01 | Microtubule-associated protein 70-2 | 1.71 |
| F6HJS7 (F6HJS7_VITVI) | CBI29537.3 | VIT_00s0527g00030.t01 | Pistil-specific extensin-like protein-like | 1.78 |
| DNA replication/Repair (n = 5) | ||||
| F6HAC8 (F6HAC8_VITVI) | CBI24290.3 | VIT_06s0009g02520.t01 | Replication factor C subunit 1-like | 1.25 |
| D7TIQ6 (D7TIQ6_VITVI) | CBI30132.3 | VIT_08s0007g05120.t01 | DNA gyrase subunit B | 0.76 |
| D7SH27 (D7SH27_VITVI) | CBI15707.3 | VIT_17s0000g00910.t01 | Histidine kinase-, DNA gyrase B-, and HSP90-like ATPase family | 1.27 |
| D7TDE2 (D7TDE2_VITVI) | CBI34931.3 | VIT_01s0127g00840.t01 | OB-fold nucleic acid binding domain containing protein | 1.33 |
| F6HSF7 (F6HSF7_VITVI) | CBI33677.3 | VIT_14s0006g03280.t01 | DNA double-strand break repair rad50 ATPase | 1.48 |
| Epigenetic regulation (n = 29) | ||||
| E0CRG0 (E0CRG0_VITVI) | CBI19114.3 | VIT_18s0001g04770.t01 | Apoptotic chromatin condensation inducer in the nucleus | 0.71 |
| A5BH86 (A5BH86_VITVI) | CBI30396.3 | VIT_08s0007g02200.t01 | High mobility group B protein 1 isoform X2 | 1.50 |
| F6HUL8 (F6HUL8_VITVI) | CBI34365.3 | VIT_02s0025g00090.t01 | RNA-binding protein C25G10.01 | 1.53 |
| D7T3I0 (D7T3I0_VITVI) | CBI25061.3 | VIT_00s0179g00340.t01 | Histone H2A.1 | 1.71 |
| F6GV41 (F6GV41_VITVI) | CBI16181.3 | VIT_06s0004g04230.t01 | Histone H2B | 1.74 |
| D7U2L4 (D7U2L4_VITVI) | CBI36980.3 | VIT_07s0005g01810.t01 | Agenet domain-containing protein | 1.87 |
| D7TCM4 (D7TCM4_VIT | CBI27882.3 | VIT_11s0016g01890.t01 | Single myb histone | 1.33 |
| D7TED8 (D7TED8_VITVI) | CBI28861.3 | VIT_12s0059g01310.t01 | SUMO protein | 1.43 |
| D7TUZ2 (D7TUZ2_VITVI) | CBI34317.3 | VIT_14s0030g00480.t01 | RNA recognition motif family protein | 1.26 |
| D7SIC8 (D7SIC8_VITVI) | CBI15238.3 | VIT_17s0000g06030.t01 | Nucleosome/chromatin assembly factor group | 1.28 |
| F6I550 (F6I550_VITVI) | CBI39729.3 | VIT_19s0015g00430.t01 | DEK domain-containing chromatin associated protein | 1.23 |
| D7T5E7 (D7T5E7_VITVI) | CBI25730.3 | VIT_00s0194g00020.t01 | DNA-directed RNA polymerases IV and V subunit 4 isoform X1 | 1.41 |
| A5BLU3 (A5BLU3_VITVI) | CBI15554.3 | VIT_17s0000g02550.t01 | Histone H1 | 1.39 |
| D7TSR3 (D7TSR3_VITVI) | CBI33535.3 | VIT_14s0006g01440.t01 | Double-stranded RNA-binding protein 4-like | 1.22 |
| E0CQU8 (E0CQU8_VITVI) | CBI18902.3 | VIT_18s0001g00660.t01 | High mobility group B protein 9 | 1.27 |
| D7U016 (D7U016_VITVI) | CBI35962.3 | VIT_09s0002g02330.t01 | Nucleosome assembly protein 1-like isoform 1 | 1.28 |
| D7U469 (D7U469_VITVI) | CBI37645.3 | VIT_04s0044g00110.t01 | High mobility group B2 protein-like isoform 1 | 1.29 |
| F6HDQ3 (F6HDQ3_VITVI) | CBI26253.3 | Not available | Suppressor of gene silencing like protein | 1.29 |
| D7U7P2 (D7U7P2_VITVI) | CBI38821.3 | VIT_15s0048g01290.t01 | Histone deacetylase complex subunit SAP18 | 1.33 |
| D7U4F5 (D7U4F5_VITVI) | CBI37552.3 | VIT_04s0044g01140.t01 | RNA-binding protein 8A | 1.37 |
| D7U5H5 (D7U5H5_VITVI) | CBI37994.3 | VIT_03s0038g00620.t01 | Zinc finger protein GIS2-like isoform 2 | 0.36 |
| D7T8P4 (D7T8P4_VITVI) | CBI26865.3 | VIT_01s0011g05360.t01 | HMG-Y-related protein A | 1.41 |
| D7UB91 (D7UB91_VITVI) | CBI40015.3 | VIT_15s0024g00620.t01 | Chromo domain protein LHP1-like heterochromatin protein 1 | 1.43 |
| F6GWG2 (F6GWG2_VITVI) | CBI17313.3 | VIT_05s0029g00130.t01 | High mobility group B protein 15-like | 1.44 |
| D7SN59 (D7SN59_VITVI) | CBI17088.3 | VIT_06s0061g01240.t01 | Histone deacetylase HDT1-like | 1.48 |
| F6HND0 (F6HND0_VITVI) | CBI31410.3 | VIT_13s0019g04940.t01 | Protein RNA-directed DNA methylation 3 isoform X1 | 1.58 |
| D7UDB2 (D7UDB2_VITVI) | CBI40727.3 | VIT_18s0122g01190.t01 | High mobility group-like isoform 1 | 1.75 |
| F6HTB7 (F6HTB7_VITVI) | CBI33920.3 | VIT_02s0012g00870.t01 | Nucleic acid binding protein | 1.89 |
| F6HIR3 (F6HIR3_VITVI) | CBI29042.3 | VIT_10s0042g00830.t01 | Lysine-specific demethylase 3B-like | 1.92 |
| Metabolism (n = 13) | ||||
| F6GSG7 (F6GSG7_VITVI) | CBI14856.3 | VIT_17s0000g10430.t01 | Glyceraldehyde-3-phosphate dehydrogenase, cytosolic | 1.95 |
| D7TGC8 (D7TGC8_VITVI) | CBI29552.3 | VIT_00s0769g00010.t01 | Peptidyl-prolyl cis-trans isomerase FKBP62 | 1.27 |
| F6H4R0 (F6H4R0_VITVI) | CBI21690.3 | VIT_19s0027g01660.t01 | Peptidyl-prolyl cis-trans isomerase CYP59 isoform X1 | 1.68 |
| D7U1R3 (D7U1R3_VITVI | CBI36679.3 | VIT_05s0102g00560.t01 | Peptidyl-prolyl cis-trans isomerase E | 2.04 |
| F6HTX9 (F6HTX9_VITVI) | CBI34281.3 | VIT_14s0030g00950.t01 | Superoxide dismutase [Cu-Zn]-like isoform 2 | 1.35 |
| F6H0A2 (F6H0A2_VITVI) | CBI19970.3 | VIT_18s0001g15570.t01 | Acetyl-CoA carboxylase carboxyltransferase subunit beta | 1.21 |
| D7FBB2 (D7FBB2_VITVI) | CBI25114.3 | VIT_16s0100g00580.t01 | Nitrogen regulatory protein P-II homolog | 1.23 |
| D7TQA5 (D7TQA5_VITVI) | CBI32625.3 | VIT_08s0040g03150.t01 | Cytosolic ascorbate peroxidase | 1.39 |
| D7SKR5 (D7SKR5_VITVI) | CBI16243.3 | VIT_06s0004g03550.t01 | L-ascorbate peroxidase 2, cytosolic | 1.39 |
| D7UDY0 (D7UDY0_VITVI) | CBI40945.3 | VIT_00s0260g00060.t01 | FK506-binding protein 2-1 | 1.41 |
| D7T674 (D7T674_VITVI) | CBI25995.3 | VIT_05s0020g00600.t01 | 1-Cys peroxiredoxin | 1.47 |
| F6HIE6 (F6HIE6_VITVI) | CBI28862.3 | VIT_12s0059g01320.t01 | O-Glycosyl hydrolases family 17 protein | 1.67 |
| D7TC92 (D7TC92_VITVI) | CBI27750.3 | VIT_11s0016g00420.t01 | Protein Red isoform 1 | 2.01 |
| mRNA-splicing/Stability/Editing (n = 42) | ||||
| D7ST85 (D7ST85_VITVI) | CBI19999.3 | VIT_12s0055g00360.t01 | Pinin/SDK/memA protein | 1.39 |
| F6HR01 (F6HR01_VITVI) | CBI32700.3 | VIT_08s0040g02300.t01 | DEAD-box ATP-dependent RNA helicase 57 | 1.45 |
| D7TUX6 (D7TUX6_VITVI) | CBI34301.3 | Not available | DCD (Development and Cell Death) domain protein | 1.46 |
| F6GUY6 (F6GUY6_VITVI) | CBI16084.3 | VIT_06s0004g05220.t01 | Serine/arginine repetitive matrix protein | 1.50 |
| D7SWX8 (D7SWX8_VITVI) | CBI21778.3 | VIT_19s0027g00590.t01 | RNA-binding protein with serine-rich domain-containing protein | 1.52 |
| A5AII6 (A5AII6_VITVI) | CBI37603.3 | VIT_04s0044g00540.t01 | Pre-mRNA-splicing factor ISY1 homolog isoform 1 | 1.63 |
| E0CRK0 (E0CRK0_VITVI) | CBI19154.3 | VIT_18s0001g05550.t01 | Splicing factor, arginine/serine-rich | 1.68 |
| F6HC22 (F6HC22_VITVI) | CBI25319.3 | VIT_13s0067g03600.t01 | Arginine/serine-rich splicing factor | 1.86 |
| D7TBV2 (D7TBV2_VITVI) | CBI28137.3 | VIT_11s0016g04610.t01 | RNA-binding protein 25 | 1.74 |
| D7TJ87 (D7TJ87_VITVI) | CBI30313.3 | VIT_08s0007g03130.t01 | Small nuclear ribonucleoprotein G | 1.74 |
| F6GSZ6 (F6GSZ6_VITVI) | CBI15706.3 | VIT_17s0000g00960.t01 | Omega-hydroxypalmitate O-feruloyl transferase | 1.76 |
| F6HZ42 (F6HZ42_VITVI) | CBI26627.3 | VIT_07s0005g00320.t01 | DEAD-box ATP-dependent RNA helicase 32 | 1.93 |
| F6GU40 (F6GU40_VITVI) | CBI16368.3 | VIT_06s0004g02220.t01 | Heterogeneous nuclear ribonucleoprotein F-like | 1.93 |
| F6GUK3 (F6GUK3_VITVI) | CBI16510.3 | VIT_06s0004g00710.t01 | SC35-like splicing factor 33 | 1.21 |
| A5AES3 (A5AES3_VITVI) | CBI24269.3 | VIT_06s0009g02770.t01 | Pre-mRNA branch site p14-like protein | 1.61 |
| F6GXF2 (F6GXF2_VITVI) | CBI17819.3 | VIT_11s0052g01130.t01 | CD2 antigen cytoplasmic tail-binding protein | 1.31 |
| F6HYI9 (F6HYI9_VITVI) | CBI36522.3 | VIT_11s0037g00130.t01 | FIP1[V]-like protein | 1.81 |
| D7T5U0 (D7T5U0_VITVI) | CBI25873.3 | VIT_00s0625g00040.t01 | Polyadenylate-binding protein 2 | 1.41 |
| F6HTT9 (F6HTT9_VITVI) | CBI34206.3 | VIT_14s0030g01680.t01 | MKI67 FHA domain-interacting nucleolar phosphoprotein | 1.27 |
| D7TU07 (D7TU07_VITVI) | CBI33922.3 | VIT_02s0012g00850.t01 | Pre-mRNA-splicing factor CWC26 | 1.29 |
| F6HYH6 (F6HYH6_VITVI) | CBI36502.3 | VIT_04s0159g00020.t01 | Polyadenylate-binding protein | 1.28 |
| D7TLV0 (D7TLV0_VITVI) | CBI31687.3 | VIT_13s0019g01060.t01 | Serine/arginine rich splicing factor | 1.32 |
| D7SJN7 (D7SJN7_VITVI) | CBI15863.3 | VIT_06s0004g07530.t01 | Spliceosomal protein | 1.35 |
| F6H257 (F6H257_VITVI) | CBI20322.3 | VIT_19s0014g02920.t01 | Pentatricopeptide repeat-containing protein | 1.28 |
| D7T3P2 (D7T3P2_VITVI) | CBI25124.3 | VIT_03s0088g00390.t01 | DnaJ homolog subfamily C member 17-like | 1.25 |
| F6GWL6 (F6GWL6_VITVI) | CBI17355.3 | VIT_04s0023g03630.t01 | Pre-mRNA-splicing factor CWC25 | 1.29 |
| F6H2X4 (F6H2X4_VITVI) | CBI20826.3 | VIT_04s0008g03130.t01 | Pre-mRNA-splicing factor CWC21-like | 1.30 |
| D7TT33 (D7TT33_VITVI) | CBI33655.3 | VIT_14s0006g02960.t01 | Poly C-binding protein | 0.36 |
| F6I0Z0 (F6I0Z0_VITVI) | CBI37849.3 | VIT_03s0038g02620.t01 | Splicing factor | 1.39 |
| F6GYT6 (F6GYT6_VITVI) | CBI18525.3 | VIT_18s0117g00150.t01 | Heterogeneous nuclear ribonucleoprotein 27C | 1.44 |
| F6HP66 (F6HP66_VITVI) | CBI31839.3 | VIT_16s0100g00450.t01 | Arginine/serine-rich-splicing factor RSP40 | 1.44 |
| D7UAL8 (D7UAL8_VITVI) | CBI39783.3 | VIT_19s0015g00980.t01 | Pre-mRNA-splicing factor cwc15 | 1.45 |
| D7TAD5 (D7TAD5_VITVI) | CBI27458.3 | VIT_01s0010g01410.t01 | RNA-binding protein-like | 1.53 |
| F6I0P5 (F6I0P5_VITVI) | CBI37715.3 | VIT_03s0038g04130.t01 | DEAD-box ATP-dependent RNA helicase 42-like | 1.56 |
| F6H177 (F6H177_VITVI) | CBI19367.3 | VIT_18s0001g08680.t01 | Pre-mRNA-processing protein 40B | 1.62 |
| F6GTQ4 (F6GTQ4_VITVI) | CBI14910.3 | VIT_17s0000g09680.t01 | 31 kDa ribonucleoprotein | 1.67 |
| F6GWX4 (F6GWX4_VITVI) | CBI17535.3 | VIT_04s0023g01580.t01 | U1 small nuclear ribonucleoprotein 70 kDa | 1.70 |
| F6HI04 (F6HI04_VITVI) | CBI28632.3 | VIT_04s0043g00270.t01 | Pre-mRNA-splicing factor 38B | 1.72 |
| F6I0K0 (F6I0K0_VITVI) | CBI37648.3 | VIT_04s0044g00080.t01 | Heterogeneous nuclear ribonucleoprotein F | 1.78 |
| D7UD56 (D7UD56_VITVI) | CBI40671.3 | VIT_11s0078g00440.t01 | U4/U6.U5 tri-snRNP-associated protein | 1.82 |
| F6HF25 (F6HF25_VITVI) | CBI27081.3 | VIT_01s0011g02820.t01 | Protein decapping 5 isoform X1 | 1.85 |
| F6HTK3 (F6HTK3_VITVI) | CBI34075.3 | VIT_03s0017g01340.t01 | Heterogeneous nuclear ribonucleoprotein Q | 2.10 |
| rRNA processing/Biogenesis (n = 13) | ||||
| D7T103 (D7T103_VITVI) | CBI24130.3 | VIT_19s0085g01090.t01 | Nuclear-pore anchor-like | 1.29 |
| E0CQ61 (E0CQ61_VITVI) | CBI19866.3 | VIT_18s0001g14320.t01 | Nucleolar protein 58 isoform X1 | 1.52 |
| F6GST5 (F6GST5_VITVI) | CBI15641.3 | VIT_17s0000g01640.t01 | RNA-metabolising metallo-beta-lactamase family protein | 1.93 |
| F6H683 (F6H683_VITVI) | CBI22501.3 | VIT_03s0091g00320.t01 | Ribosomal RNA assembly protein mis3-like | 0.72 |
| F6HLD3 (F6HLD3_VITVI) | CBI30568.3 | VIT_08s0007g00190.t01 | H/ACA ribonucleoprotein complex subunit 4 | 1.45 |
| D7T1S3 (D7T1S3_VITVI) | CBI24453.3 | VIT_00s0264g00120.t01 | Scaffold attachment factor B1 | 1.91 |
| C5DB53 (C5DB53_VITVI) | CBI31135.3 | VIT_08s0056g00160.t01 | U3 small nucleolar RNA-associated protein 11 | 1.34 |
| F6GZQ7 (F6GZQ7_VITVI) | CBI16359.3 | VIT_18s0001g13560.t01 | Midasin | 1.21 |
| F6HF03 (F6HF03_VITVI) | CBI27323.3 | VIT_01s0011g00070.t01 | Nucleolar protein 14-like | 1.50 |
| D7STQ8 (D7STQ8_VITVI) | CBI20657.3 | VIT_04s0008g01200.t01 | Translation machinery-associated protein 22 isoform 2 | 0.33 |
| F6I6B2 (F6I6B2_VITVI) | CBI40495.3 | VIT_15s0046g01120.t01 | Ribosome biogenesis regulatory protein homolog | 1.58 |
| D7U276 (D7U276_VITVI) | CBI36842.3 | VIT_07s0005g00270.t01 | Nucleolar protein 16 involved in ribosome biogenesis | 1.90 |
| F6GXL7 (F6GXL7_VITVI) | CBI17936.3 | VIT_07s0141g00380.t01 | U3 small nucleolar RNA-associated protein-like protein | 2.00 |
| Stress responses (n = 13) | ||||
| F6HS56 (F6HS56_VITVI) | CBI33350.3 | VIT_05s0051g00650.t01 | Voltage-gated potassium channel subunit beta | 0.72 |
| E0CUG6 (E0CUG6_VITVI) | CBI22747.3 | VIT_16s0050g00140.t01 | Metal ion binding protein | 1.63 |
| F6GY60 (F6GY60_VITVI) | CBI18236.3 | VIT_18s0072g00160.t01 | Peroxidase 12-like | 1.34 |
| D7TUZ6 (D7TUZ6_VITVI) | CBI34321.3 | VIT_14s0030g00430.t01 | Prefoldin chaperone subunit family protein | 1.21 |
| E0CRL1 (E0CRL1_VITVI) | CBI19165.3 | VIT_18s0001g05720.t01 | 14-3-3 protein 7 | 1.27 |
| F6H0X3 (F6H0X3_VITVI) | CBI19195.3 | VIT_18s0001g06330.t01 | 14-3-3 protein | 1.49 |
| F6H824 (F6H824_VITVI) | CBI23432.3 | VIT_00s0250g00040.t01 | DNA-binding protein | 1.21 |
| D7SU28 (D7SU28_VITVI) | CBI20777.3 | VIT_04s0008g02590.t01 | Selenium binding protein | 1.22 |
| F6HEA6 (F6HEA6_VITVI) | CBI26439.3 | VIT_16s0039g01020.t01 | Adenylate cyclase, terminal-differentiation specific | 1.30 |
| D7TIR0 (D7TIR0_VITVI) | CBI30136.3 | Not available | Arginine/serine-rich coiled-coil protein 2 isoform X2 | 1.43 |
| F6H1I0 (F6H1I0_VITVI) | CBI19683.3 | VIT_18s0001g12350.t01 | Protein ESSENTIAL FOR POTEXVIRUS ACCUMULATION X1 | 1.59 |
| F6H2Z1 (F6H2Z1_VITVI) | CBI20901.3 | Not available | Dehydration-responsive protein RD22 | 1.93 |
| F6HE42 (F6HE42_VITVI) | CBI26016.3 | VIT_05s0020g00840.t01 | Late embryogenesis abundant protein D-29 | 4.17 |
| Transcriptional regulation (n = 20) | ||||
| F6H7R2 (F6H7R2_VITVI) | CBI23284.3 | VIT_07s0197g00070.t01 | Upstream activation factor subunit spp27-like | 1.59 |
| D7SIK8 (D7SIK8_VITVI) | CBI15319.3 | VIT_17s0000g05190.t01 | Zinc finger CCCH domain-containing protein | 1.64 |
| D7TCU3 (D7TCU3_VITVI) | CBI28316.3 | VIT_06s0080g00460.t01 | Nuclear transcription factor Y subunit B-8 | 1.71 |
| D7TDY1 (D7TDY1_VITVI) | CBI28704.3 | VIT_07s0151g00910.t01 | NF-kappa-B-activating protein | 1.21 |
| F6HSW0 (F6HSW0_VITVI) | CBI33736.3 | VIT_07s0129g00610.t01 | FRIGIDA-like isoform 2 | 1.28 |
| F6I111 (F6I111_VITVI) | CBI37898.3 | VIT_03s0038g02130.t01 | Cold-shock DNA binding protein | 1.47 |
| D7UDF0 (D7UDF0_VITVI) | CBI40765.3 | Not available | AT-hook protein 1 | 1.36 |
| D7SK51 (D7SK51_VITVI) | CBI16027.3 | VIT_06s0004g05830.t01 | DNA-directed RNA polymerases I and III subunit RPAC2 isoform 1 | 1.25 |
| F6HZB5 (F6HZB5_VITVI) | CBI36973.3 | VIT_07s0005g01740.t01 | Zinc knuckle (CCHC-type) family protein | 1.37 |
| F6HIW1 (F6HIW1_VITVI) | CBI29150.3 | VIT_13s0047g00310.t01 | Serrate RNA effector molecule-like | 1.37 |
| E0CNQ9 (E0CNQ9_VITVI) | CBI19287.3 | VIT_18s0001g07750.t01 | Neuroguidin-like | 1.38 |
| F6HAX1 (F6HAX1_VITVI) | CBI24668.3 | VIT_05s0094g00440.t01 | Sas10/U3 ribonucleoprotein family protein | 1.42 |
| D7SII5 (D7SII5_VITVI) | CBI15296.3 | VIT_17s0000g05450.t01 | Early flowering 5 protein | 1.43 |
| D7TZU6 (D7TZU6_VITVI) | CBI35892.3 | VIT_09s0002g01530.t01 | GBF-interacting protein 1-like isoform X1 | 1.52 |
| F6HH48 (F6HH48_VITVI) | CBI28116.3 | VIT_11s0016g04390.t01 | Nucleolar protein dao-5-like | 1.54 |
| F6HFZ8 (F6HFZ8_VITVI) | CBI27460.3 | VIT_01s0010g01440.t01 | Transcription elongation regulator 1-like | 1.57 |
| D7TTQ2 (D7TTQ2_VITVI) | CBI33817.3 | VIT_02s0012g02250.t01 | Transcription factor HBP-1a | 1.69 |
| F6I758 (F6I758_VITVI) | CBI40894.3 | VIT_13s0175g00120.t01 | ABSCISIC ACID-INSENSITIVE 5-like protein 2-like isoform 1 | 1.75 |
| F6HLJ6 (F6HLJ6_VITVI) | CBI30026.3 | VIT_08s0007g06400.t01 | Zinc finger CCCH domain-containing protein 14-like isoform 1 | 1.83 |
| D7SIC5 (D7SIC5_VITVI) | CBI15235.3 | VIT_17s0000g06060.t01 | Activating signal cointegrator 1 | 2.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atanassov, H.; Parrilla, J.; Artault, C.; Verbeke, J.; Schneider, T.; Grossmann, J.; Roschitzki, B.; Atanassova, R. Grape ASR-Silencing Sways Nuclear Proteome, Histone Marks and Interplay of Intrinsically Disordered Proteins. Int. J. Mol. Sci. 2022, 23, 1537. https://doi.org/10.3390/ijms23031537
Atanassov H, Parrilla J, Artault C, Verbeke J, Schneider T, Grossmann J, Roschitzki B, Atanassova R. Grape ASR-Silencing Sways Nuclear Proteome, Histone Marks and Interplay of Intrinsically Disordered Proteins. International Journal of Molecular Sciences. 2022; 23(3):1537. https://doi.org/10.3390/ijms23031537
Chicago/Turabian StyleAtanassov, Hristo, Jonathan Parrilla, Caroline Artault, Jérémy Verbeke, Thomas Schneider, Jonas Grossmann, Bernd Roschitzki, and Rossitza Atanassova. 2022. "Grape ASR-Silencing Sways Nuclear Proteome, Histone Marks and Interplay of Intrinsically Disordered Proteins" International Journal of Molecular Sciences 23, no. 3: 1537. https://doi.org/10.3390/ijms23031537