This site is best viewed on Microsoft Internet Explorer 4.0 or higher
with screen set to 1024 X 768 pixels, 24 bit …16 million colors.
Set … View/Text Size … to Meduim
Click on thumbnails to view larger images.
——————————————————————————–
For more than twenty years, I have been studying the image generating properties of reflective spheres stacked in 52 degree angle pyramids. The 52 (51.827) degree angle slope of the sides of The Great Pyramid in Cairo, Egypt embodies the Golden Mean which is the ratio that is used in Nature to generate growth patterns in space. Sacred Geometry studies such primal systems which reveal the unity of the cosmos by representing the relationships between numbers geometrically. The Vesica Piscis is one of the most fundamental geometrical forms of this ancient discipline and it reveals the relationship between the The Great Pyramid and the 2 dimensional expansion of a circle of one unit radius R as shown in Figure 1. This relationship is more completely described in The New View Over Atlantis by John Michell published by Thames and Hudson.
Figure 1 Vesica Piscis in 2 Dimensions
In the early 1970s, I became very interested in the three dimensional representation of this geometry and I visualized this as a three dimensional pyramid inside two intersecting spheres shown in Figure 2.
Figure 2 Vesica Piscis in 3 Dimensions
In an effort to visualize these 3D relationships in yet another way, I stacked reflective Christmas Tree balls in an inverted pyramid shell. I discovered that the patterns of multiple reflections created on the interior surfaces of reflective spheres stacked at this angle produce images that relate to the human form as photographed and shown in Figure 3 and Figure 4.
I made many pyramidal configurations of reflective spheres with different light sources and I photographed the patterns on the interior surfaces from many points of view. Another is shown here in Figure 5.
Then in 1977, I discovered one stacking structure and viewpoint that produced a very convincing image of an archetypal human face. This structure of 10 spheres (2 5-ball pyramids) forming a cluster is shown in Figure 6.
The face image is shown in the lower third of the pattern on one sphere inside this 10 sphere cluster and is shown in Figure 7.
Figure 7 When I made this photograph, the structure was enclosed in a mirrored pyramid.
Later, I realized that the most natural structure for enclosing would be another 10 sphere, 2 pyramid structure that would totally enclose a smaller but similar cluster. I worked out the math to find that by multiplying the inner sphere’s diameters by Pi gives the dimension for the outer sphere’s diameters as shown in Figure 8.
Figure 8 One sphere is removed from the outer cluster to reveal the inner cluster. However, the inner cluster must be upside down with respect to the outer cluster to fit inside.
The expansion by Pi reinforced my suspicion that this 10 sphere cluster is a fundamental unit that is linked to the properties of three dimensional space.
Close-packed reflective spheres clustered in this concentric shell structure produce an optical distribution network that links the Golden Mean and Pi. The Golden Mean is expressed in the 52 degree angle pyramid structure and Pi is expressed in the ratio of the diameter to the circumference of each sphere of course; but it is also expressed in the ratio of the sizes of spheres in the 10 spheres within 10 spheres concentric shell structure that I discovered. This concentric shell structure could continue to expand with many shells and still retain the same ratio between shells. It was not until 1991 that I was able to build and photograph the images inside a ten-within-ten (2 shell) structure. I used 10 – 6″ diameter spheres and 10 – 19″ diameter hemispheres. The structure is shown here in Figure 9.
The photographs that I made from this 10-within-10 sphere structure revealed the inherent limitations of photography for this work. The final images were not what I was seeing with my eyes while directly viewing the interior of the structure. But this approach did reveal a more complete face form and I also realized that I would need at least 10 more spheres (about 60″ in diameter at $1500 each) to complete the enclosure and remove the remaining gaps in the images. Also, the lighting system was limited to the exterior and it was very difficult to control the positioning, color and brightness. So, in 1992, I purchased a computer to model these structures with ray-tracing software which enabled me to investigate more thoroughly the relationship between this cluster geometry and the archetypal images generated therein. During this tour into cyberspace, I could take a camera into the sacred chamber central to the concentric shells of reflective spheres which are simulated by a computer program (Real 3D by Realsoft Oy, Finland) that most accurately renders the effects of real-world light sources and records the patterns generated by multiple reflections on metal surfaces. With this method of investigation, I am able to more conveniently control the many variables which led to these discoveries and conclusions:
1.) At least 40 spheres (4-10 ball shells) are necessary to enclose the central area and fill in the gaps in the images.
2.) Most of the lights should be point sources placed at the points of contact between the innermost spheres of the structure; although additional symmetrically-paired, point-lights are necessary in the central area.
3.) Some of the innermost spheres are reduced in size and they can float within certain areas in the central space. Figure 12 shows typical positions and sizes.
4.) The camera position and field of view as shown in Figure 12 produces the most convincing image of the human form.
5.) The image of this artificial anthropoid that is produced in these structures can be animated when the positions and sizes of certain spheres are modulated as shown in the three animations listed below.
animation 1 .8 MB … estimated download time at 56k … 3.5 min.
animation 2 2.4 MB … estimated download time at 56k … 10 min.
Figure 13 Human Form From Sacred Geometry
The computer image in Figure 13 was made from the camera position and field of view shown in Figure 12.
Figure 14 is from the same camera position (shown in Figure 12) but the lens set at a very wide angle.
Figure 16 is a stereo image and it shows the interior of the cluster in 3D. Stare through the images with your eyes focused at a distant point and the two images will turn into three images and the center image will appear in 3D.
In 1996, I produced an animation Sacred Spaces (6 minutes) which has been screened in many national and international film festivals and it has won some awards (see resume). I have also produced Flesh Tones (5.5 Minutes) another animation completed in 1997 and I have produced many high resolution images for prints/slides (some examples are shown in Figures 13 through 36) which I have presented with the video animations at lecture presentations. These images are stills from the animations and they demonstrate the variety of image generating that is inherent in this system. I have concentrated on exhibiting the videotape documentation of my work thus far because it is the most portable presentation format that describes this research most completely.
This interdisciplinary research has taken me into many related areas of study. Geometry, Optics, Ancient History of Art and Religion, Computer Imaging, Photography, Animation, Graphic Art and of course Sculpture are the major connecting disciplines that have contributed to this work.
I feel that I have rediscovered some of what was a highly developed understanding of Mankind’s relationship to the Universe and this knowledge was utilized and documented in the geometry of ancient structures. Sacred Geometry, the study of the unity of the cosmos, demonstrates relationships between Number and Space and the Human Form. It was used in the construction of ancient glyphs and monuments thereby preserving the knowledge of these principles of Natural Law for future generations. This construction of reflective spheres may embody the technology that produced the animated images of the deities in the temples of antiquity. The Tree of Life which is a graphic representation of the interaction between cosmic forces is shown in Figure 22. It is found in many ancient texts of the Kabbalah.
The construction of this structure of clustered metal reflective spheres (offering bowls) is well within the capability of many ancient cultures and with the addition of a few glass lenses, these images could be projected onto walls or into smoke. Perhaps there is some Truth behind the smoke and mirrors of Ancient Religion … perhaps it is geometry … Sacred Geometry.
This research which is documented in four computer animations Sacred Spaces, Flesh Tones, Our Mothers and Sacred Spaces 2, in color computer prints, and in color slides has given me new insight into the motives that may have inspired the construction of The Great Pyramid.
1998 – 2000 update
The images shown in Figures 24 – 35 were made with 50 spheres and 144 point-light sources. Each of these images was made with unique brightness, color and value settings for various groupings of lights.
InFigure32, the camera is aimed at the sphere opposite the face shown inFigure 31.Figure 12 shows the camera position for Figure 31. The image in Figure 32 was formed when the camera position was rotated 180 degrees around the vertical axis shown in Figurre 12 and zoomed in. The face image (a child?) in Figure 32 is much smaller than the face (mother?) in Figure 31.
Figure35This is a stereo 3D image that requires shutter glasses to view and your monitor must be set to interlace mode.
Figure 36 Stereo Image for Cross-Eyed Viewing
With your monitor at arm’s length away, focus on a point 6 inches in front of your nose (put your index finger 6 inches in front of your nose and focus on it). You will see a third image in 3D between the two images on the monitor (at the tip of your finger). Shift your attention from your finger to this third/middle image which will appear in 3D.
———————————————————————————
The cross-eyed viewing method is perhaps the most effective way to put the viewer inside the cluster to see the human image as it would exist in 3D from the cameras position as seen in Figure 12. There are many more identifiable images in this clustering geometry viewed from this position and from other camera positions and even more images with other color settings for the point lights.
Figure 37 Rods connecting centers of nearest neighbors in 3 shells
I’m now very curious about the relationship between Sacred Geometry and Sacred Music and the Human Form. Number relates to all that science measures by virtue of the way that 3D Space is defined. Number is also used to measure Time … and, as Pythagoras observed, Music is a manifestation of Number in Time. The distribution of sounds i.e. amplitude and frequency, may well find an idealized model in the 10 within 10 sphere, space-filling, close-packing geometric system. The representation of this geometry with sticks or strings or rods as shown in Figure 37 (in which the centers of nearest neighbor spheres in three shells are joined) may represent the ideal space-filling matrix of linear oscillating elements. It may also be used to define spatially distributed, hierarchical, cellular arrays.
————————————————————————
The camera positions on the symmetric planes within the cluster produce a bilateral symmetry that we identify with animal and human form. Naturally, we would expect this symmetry in the idealized images of higher life forms. The multiplicity of idealized beings in this cluster of reflective spheres suggests the presence of The Company of the Gods as described in many ancient Egyptian texts. It seems that the face images occur on a vertical plane linking the centers of spheres. There seem to be faces facing faces and faces within faces throughout this reflective environment. My guess is that the sacred part of what I have discovered is a result of the way the deity put the higher life forms in three dimensions.
This cluster geometry may have other properties that would be useful for spatial organization. The nesting of 10 reflective spheres within 10 reflective spheres geometry produces a distribution system that could be used for processing of optical information between the interior to the exterior of the structure. Because of the spatial distribution of the reflective spheres in two concentric shells of ten each, optical information must be reflected and diverged in order to enter or exit the system with the exception of a few radially arrayed directions. The system becomes a more selective filter of optical information as more shells are added to the structure. This inside-to-outside transformation/translation should have many practical applications in pattern recognition tasks. For example, any point source of coherent light (laser light) anywhere outside the two-shell cluster will produce a unique light distribution pattern on the inside as viewed from the center area of the cluster. This pattern could be recorded in a holographic medium and the exterior point source could then be reconstructed using conventional holographic means.
Is Number (Geometry and Time) the link between Art, Science and Religious experience? The language of number is perhaps the most convincing form of expression between humans and between humans and the Gods. We think and imagine in visual forms. Einstein constructed his mathematics based on mental images. He said that he would first try to visualize a space/time image and then mathematize it. We use mental images to construct possible scenarios of the future so that we don’t have to live out each one in “reality”. Words and pictures and mathematical formulas are ways to document, test, realize, and communicate these visions. Although there is seldom a need to mathematize images, we sense that it would be possible. We know that we could count and number the grains of sand on the beach. The geometry of our visions is what makes them real to us and it allows us to mentally work on them and to integrate them convincingly into our life here in 3D.
I’m not quite sure how the physical human form fits into the grand scheme of things but it does seem to be a result of the space-filling, spherical, close-packing geometric system that I’ve discovered and it is indeed “Sacred Geometry” by virtue of the definition God gave to three dimensions. I don’t think this geometric system is the matrix for all life systems but I think it can serve as a model for the interactions between the various dimensional realities in which we are immersed. It may also guide us in our attempt to develop new sciences and technologies that utilize the forces that operate in the regions that we now call consider paranormal.
According to contemporary Superstring Theory as described by Dr. Michio Kaku in his many recent writings, the mathematics that most appropriately describes the forces of nature requires an expression in ten dimensions. Einstein tried to describe the forces of nature in the mathematics of 3D and Time and found that the formulas were not broad enough to include all of the forces. We can perceive 3D and Time. The other 6D in Superstrings are hidden from our normal senses due to their incredibly small size according to Dr. Kaku. Our instincts inform us that there are more than 3 Dimensions and Time in the universe and the possiblity that something else exists mathematically beyond our perceptual horizon drives my curiosities about our possible links to these worlds. The new science of parapsychology has discovered many ways that humans can perceive by means beyond the physical senses and it has found that there are some people that are more capable of extrasensory perception than others. It may well be that our only contacts with this duality of nature are through numbers and mathematics on one hand or through dreams, remote viewing, telepathy, Ouija boards and tarot cards on the other. This would certainly confirm the existence of a God with a sense of humor
I am interested in any information relating this technology to ancient religious traditions. Any references that you send will certainly be appreciated and I will certainly credit any references in future publications.
A more detailed version of this research is now available on CD-ROM and it includes the 10 minute animation Sacred Spaces 2 in streaming format.
I am also making available unlimited editions of selected images on this website which will be printed in very high resolution on archival paper with archival inks.
For more specific information about the availability, formats, sizes and pricing of the prints and CD relating to this work you may contact me at:
Code Biology, Bio-Semiotics, and Relational Biology
Key Terms
Biosemiotics
Anticipatory Systems
Code biology
Relational biology
C.S. Peirce
T. Sebeok
Jesper Hoffmeyer
Marcello Barbieri
Robert Rosen
Rom Harré
F Schelling
Habits
Pratibha
Innate Ability
Archetypes
Talent
Character
Virtues
Caste System
Astrology
Invariance
Regularities
Periodicities
Sapta Rishis
Evolution
Development
Biology
Codes
Meaning
Culture
Nature
Archetypes and Code Biology
Source: Archetypes and code biology
As a clinical psychologist, I observe stereotyped formulas of behavior in action every day in the consulting room, despite differences in age, race, or culture; they present themselves as codified rules or typical modes of behavior in archetypical situations. Such circumstances coincide with what C.G. Jung defended: the existence of archetypes stored in an inherited/phylogenetic repository, which he called the collective unconscious – somewhat similar to the notion of an ethogram, as shown by ethology. Psychologists can use a perspective to facilitate understanding the phenomenon: the code biology perspective (Barbieri 2014). This approach can help us recognize how these phenomenological events have an ontological reality based not only on the existence of organic information but also on the existence of organic meaning.
We are not a tabula rasa (Wilson 2000): despite the explosive diversification of the brain and the emergence of conscience and intentionality, we observe the conservation of basic instincts and emotions (Ekman 2004; Damasio 2010) not only in humans but in all mammals and other living beings; we refer to the neural activity on which the discrimination behavior is based, i.e., the neural codes. The conservation of these fundamental set-of-rules or conventions suggests that one or more neural codes have been highly conserved and serves as an interpretive basis for what happens to the living being who owns them (Barbieri 2003). Thus, archetypes’ phenomenological reality can be understood not as something metaphorical but as an ontological (phylogenetic) fact (Goodwyn 2019).
Furthermore, epigenetic regulation theories present the possibility that the biomolecular process incorporates elements of the context where it takes place; something fundamental to understand our concept – the archetype presents itself as the mnesic remnant of the behavioral history of individuals who preceded us on the evolutionary scale. In short: brains are optimized for processing ethologically relevant sensory signals (Clemens et al., 2015).
From the perspective of the corporeal mind (Searle 2002), in this paper, we will show the parallels between code biology and the concept of the archetype, as Jung defended it and as it appears in clinical practice.
Source: Code Biology 3: the study of all Codes of Life
Editorial Overview of the third special issue in code biology
Introduction This third special issue in Code Biology is a collection of highly different papers and their differences have two main causes. The first, the most obvious, is that Code Biology is the study of all codes that exist in living systems and the diversity of the papers is a direct consequence of the diversity of the codes. The second source of diversity is the existence of different theories. More precisely, the original theory that gave origin to Code Biology has been followed by a number of extended theories that now coexist with the original one. In Code Biology, in other words, there is pluralism but there has also been a beginning, and it is important to be clear about this starting point. The original theory of Code Biology is characterized by ideas that make it different from four major theoretical frameworks:
[1] The original theory of Code Biology is different from the Modern Synthesis for two reasons. The first is the idea that evolution took place by natural selection and by natural conventions and these mechanisms are fundamentally different because natural selection is based on copying and natural conventions are based on coding. The second is the idea that the cell is not a biological computer made of genotype and phenotype but a trinity of genotype, phenotype and ribotype, where the ribotype is the ribo nucleoprotein system of the cell that functions as the codemaker of the genetic code (Barbieri 1981, 1985, 2003).
[2] The original theory of Code Biology maintains that the fundamental process of life is not autopoiesis but codepoiesis (Barbieri 2012). Autopoiesis requires biological specificity and specificity comes from the genetic code, so the ancestral systems that came before that code could not have been autopoietic systems. Those ancestral systems, on the other hand, were engaged in the evolution of the genetic code and were therefore codepoietic systems. Autopoiesis, furthermore, is most evident in bacteria and bacteria have not increased their complexity and have not evolved new codes for billions of years after their appearance on Earth. It was the eukaryotes that became increasingly complex and that evolved new codes, which suggests a deep link between codes and complexity, and in particular between the origin of new codes and the origin of the great novelties of macroevolution (Barbieri 2015, 2016, 2017, 2020). Codepoiesis, on the other hand, is necessarily implemented by mechanisms, and according to the original theory of Code Biology the major mechanism that fuelled the evolution of the genetic code was the process of ambiguity reduction (Barbieri 2019a).
[3] The original theory of Code Biology is different from Biosemiotics because it claims that the Peircean processes of interpretation and abduction take place in the brain but not in the cell (Barbieri 2014,2018).
[4] The original theory of Code Biology is different from the Relational Biology of Robert Rosen because it assumes that the process of anticipation takes place in the brain but not in the cell (Barbieri 2019b).
There are, in conclusion, four key ideas in the original theory of Code Biology:
[a] Evolution took place by natural selection and by natural conventions. [b] The cell is a trinity of genotype, phenotype and ribotype. [c] The fundamental process of life is codepoiesis, not autopoiesis. [d] Ambiguity reduction was the major evolutionary mechanism of the genetic code.
The extended theories of Code Biology differ from the original theory either because they introduce new concepts or because they reformulate some of the original concepts.
[1] The first extended theory appeared when Stefan Kühn and Jan-Hendrik Hofmeyr (2014) proposed an extended definition of code, a definition where signs and meanings can be not only molecules but also biological processes. More precisely, Kühn and Hofmeyr showed that the histone code is a mapping where the signs are the marks produced on histones by acetylation or methylation processes and their meanings are the activation or the repression of particular genes.
[2] A second extended theory of Code Biology has been proposed in this issue by Julie Heng and Henry Heng with the idea that the adaptors of a biological code can be “information flows”. More precisely, Heng and Heng point out that in addition to the codes that produce the components of a system there are also codes that organize those components into a working whole. The code that is used to make bricks, for example, is different from the code that is used to construct a building from those bricks. The genetic code is a code that makes bricks, i.e., proteins, but in order to arrange proteins into a living system we need an architectural code that Heng and Heng call “karyotype code”.
[3] A third extended theory is presented in this issue by Omar Paredes and colleagues on the grounds that the original theory of Code Biology “raises the illusion that information has only an upward direction … whereas the current overview of cellular dynamics … illustrates that information flows freely upward and downward”. In order to overcome this limitation, the authors propose “a novel category of organic codes, the metacode”, which is defined as “an informational structure that handles the continuum of the information flow in biological systems”.
The extended theories, in short, are a reality and their existence is a testimony that there is genuine pluralism in Code Biology. The goal of this special issue, on the other hand, is to give a bird’s-eye view of the present status of Code Biology and to this purpose it has been divided into four parts, each of which is going to be illustrated in the rest of this editorial with brief presentations of its papers
Origin and evolution of the genetic code: the universal enigma
Eugene V. Koonin* and Artem S. Novozhilov National Center for Biotechnology Information, National Library of Medicine, National Institutes of Health, Bethesda, MD 20894
IUBMB Life. 2009 February ; 61(2): 99–111. doi:10.1002/iub.146.
I have studied complex systems science from following teachers:
Yaneer Bar Yam of NECSI: Physical, Biological, and Social Systems
Scott Page of University of Michigan: Model Thinking: Complex Systems in Economics and Social/Political Sciences.
Melanie Mitchell of Santa Fe Institute: Introduction to Complex Systems
Michael Kearns of University of Pennsylvania: Networked Life: Science of Networks
Ravi Iyenger of Mount Sinai Icahn School of Medicine: Introduction to Systems Biology
Perry Mehrling of Columbia University/INET: Economics of Money and Banking
John Sterman of MIT: Business Dynamics: Using Systems Dynamics
Key Terms
Complex Systems
Networks
Interaction
Agents
Bottom Up Modeling
Agent Based Modeling
Netlogo
New England Complex Systems Institute (NECSI)
Santa Fe Institute (SFI)
Yaneer Bar Yam
Scott Page
Melanie Mitchell
Network Science
Dynamical Systems
Cellular Automata
Artifical Life (A-Life)
Anti Fragility
Agility
Learning
Adaptability
Resilience
Synchronization
Mark Newman
Albert Laszlo Barabasi
John Sterman
Small World Networks
Scale Free Networks
Hierarchy
Boundaries
Stephen Wolfram
Stuart Kaufman
Increasing Retuns
Path Dependence
J Doyne Farmer
W. Brian Arthur
Duncan Watts
Eugene Stanley
Six Degrees of Separation
Emergence
Non Linear Dynamics
Competition and Cooperation
What are Complex Systems?
Source: A Brief History of Systems Science, Chaos and Complexity
Key Topics in Complex Systems
Sources: Complex Systems: A Survey
Lattices and Networks
Dynamical Systems
Discrete Systems and Cellular Automata
Scaling and Criticality
Adaptation and Game Theory
Information Theory
Computational Complexity
Agent based Modeling
Chaos and Fractals
Spontaneous Order and Synchronization
Complex Systems
The original journal devoted to the science, mathematics and engineering of systems with simple components but complex overall behavior; publishes high-quality articles that focus on, but are not limited to, the following areas:
Dynamic, topological and algebraic aspects of cellular automata and discrete dynamical systems
Complex systems and complexity theory
Algorithmic complexity and information theory
Emergent properties of dynamical systems
Formal languages, grammars and automata
Algorithmic information dynamics
Symbolic dynamics and connections to continuous systems
Tilings, rewriting and substitution systems
Computability theory
Synchronous versus asynchronous models
Applications of automata to areas such as machine and deep learning, physics, biology, social sciences and others
Source: A Brief History of Systems Science, Chaos and Complexity
History of Systems and Complex Systems
Source: A Brief History of Systems Science, Chaos and Complexity
Foundations of Complex-system Theories: In Economics, Evolutionary Biology …
By Sunny Y. Auyang
From simplistic to complex systems in economics
John Foster*
Cambridge Journal of Economics 2005, 29, 873–892
doi:10.1093/cje/bei083
Complex Systems: A Survey
M. E. J. Newman
Department of Physics, University of Michigan, Ann Arbor, MI 48109 and Center for the Study of Complex Systems, University of Michigan, Ann Arbor, MI 48109
Yang-YuLiu1,2 andAlbert-L ́aszl ́oBarab ́asi3,2,4,5 1Channing Division of Network Medicine, Brigham and Women’s Hospital, Harvard Medical School,
Boston, Massachusetts 02115, USA 2Center for Cancer Systems Biology, Dana-Farber Cancer Institute, Boston, Massachusetts 02115,
USA 3Center for Complex Network Research and Departments of Physics, Computer Science and Biology, Northeastern University, Boston, Massachusetts 02115, USA 4Department of Medicine, Brigham and Women’s Hospital, Harvard Medical School, Boston, Massachusetts 02115, USA 5Center for Network Science, Central European University, Budapest 1052, Hungary
a Complex Systems Theory Department, Institute of Nuclear Physics, Polish Academy of Sciences, PL–31-342 Kraków, Poland b Institute of Computer Science, Faculty of Physics, Mathematics and Computer Science, Cracow University of Technology, PL–31-155 Kraków, Poland
SENDIL K. ETHIRAJ The Wharton School of Business 3620 Locust Walk, Suite 2000 University of Pennsylvania, Philadelphia PA 19104 Ph. 215-898 1231 Email: sethiraj@wharton.upenn.edu
Complex Systems—A New Paradigm for the Integrative Study of Management, Physical, and Technological Systems
Luis A. Nunes Amaral Department of Chemical and Biological Engineering, McCormick School of Engineering, Northwestern University, Evanston, Illinois 60208, amaral@northwestern.edu
Brian Uzzi Department of Management and Organizations, Kellogg School of Management, Northwestern University, Evanston, Illinois 60208, uzzi@northwestern.edu
MANAGEMENT SCIENCE
Vol. 53, No. 7, July 2007, pp. 1033–1035 issn 0025-1909 eissn 1526-5501 07 5307 1033
THE ONTOLOGY OF COMPLEX SYSTEMS: Levels of Organization, Perspectives, and Causal Thickets*
(Canadian Journal of Philosophy, supp. vol #20, 1994, ed. Mohan Matthen and Robert Ware, University of Calgary Press, 207-274). by William C. Wimsatt Department of Philosophy University of Chicago January 4, 1994 wwim@midway.uchicago.edu
Cities as Complex Systems: Scaling, Interactions, Networks, Dynamics and Urban Morphologies
ISSN 1467-1298
Michael Batty
Centre for Advanced Spatial Analysis, University College London, 1-19 Torrington Place, London WC1E 6BT, UK Email: m.batty@ucl.ac.uk, Web: www.casa.ucl.ac.uk
The Encyclopedia of Complexity & System Science, Springer, Berlin, DE, forthcoming 2008. Date of this paper: February 25, 2008.
Scale invariance and universality: organizing principles in complex systems
H.E. Stanleya;∗, L.A.N. Amarala , P. Gopikrishnana , P.Ch. Ivanova , T.H. Keittb , V. Pleroua
aCenter for Polymer Studies and Department of Physics, Boston University, Boston, MA 02215, USA bNational Center for Ecological Analysis and Synthesis, 735 State Street, Suite 300, Santa Barbara, CA 93101, USA
aCentre for Advanced Spatial Analysis, University College London, 1 to 19 Torrington Place, London WC1E 6BT, UK bDepartment of Geography, University of Utah, 260 S. Central Campus Dr., Rm. 270, Salt Lake City, UT 84112-9155, USA
John D. Sterman Jay W. Forrester Professor of Management and Professor of Engineering Systems Sloan School of Management Massachusetts Institute of Technology 30 Wadsworth Street, E53-351 Cambridge MA 02142 617.253.1951 jsterman@mit.edu web.mit.edu/jsterman/www
Francisco A. Rodriguesa, Thomas K. DM. Peronb,c,∗, Peng Jic,d,∗, Jürgen Kurthsc,d,e,f
aDepartamento de Matemática Aplicada e Estatística, Instituto de Ciências Matemáticas e de Computação, Universidade de São Paulo, Caixa Postal 668, 13560-970 São Carlos, São Paulo, Brazil bInstituto de Física de São Carlos, Universidade de São Paulo, Caixa Postal 369, 13560-970, São Carlos, São Paulo, Brazil cPotsdam Institute for Climate Impact Research (PIK), 14473 Potsdam, Germany dDepartment of Physics, Humboldt University, 12489 Berlin, Germany eInstitute for Complex Systems and Mathematical Biology, University of Aberdeen, Aberdeen AB24 3UE, United Kingdom fDepartment of Control Theory, Nizhny Novgorod State University, Gagarin Avenue 23, Nizhny Novgorod 606950, Russia
Uncovering the overlapping community structure of complex networks in nature and society
Gergely Palla†‡, Imre Dere ́nyi‡, Ille ́s Farkas†, and Tama ́s Vicsek†‡ †Biological Physics Research Group of HAS, Pa ́zma ́ny P. stny. 1A, H-1117 Budapest, Hungary,
‡Dept. of Biological Physics, Eo ̈tvo ̈s University, Pa ́zma ́ny P. stny. 1A, H-1117 Budapest, Hungary.
In a future post, I will research structural colors.
Key Words
Color Change in Biology
Color Change using Technology
Smart Pigments
Thermochromic property
Photochromic property
Piezochromic property
Solvatochromic property
Chimiochromic property
Electrochromism
Smart Textiles
Smart Plastics
Smart Paper
Smart Inks
Smart Food Packaging
Color Science
Material Science
Color Fading
Color Fastness
Color Metamerism
Chromatophores
Iridophores
Leucophores
Chlorophyll
Anthrocyanins
Flavonols
Flavonoids
Color Change and Technology
Chromic phenomena in dyes and pigments
Some of the major companies are
LCR Hallcrest LLC
Hali Pigment Co. Ltd
Chromatic Technologies Inc.
QCR Solutions Corp.
OliKrom
SFXC
MICI
RPM International Inc.
Good Life Innovations Ltd
FX Pigments Pvt. Ltd
Smarol Industry Co. Ltd
Kolortek Co. Ltd
Kolorjet Chemicals Pvt. Ltd
Colourchange
Source: OliKrom
Smart Hybrid Pigments
The solutions developed by OliKrom involve a new generation of hybrid pigments that combines the proven strength of the metal ions and the flexibility of the molecular material. The change in the structure allow to control the color change as a function of :
Temperature (thermochromic property),
Light (photochromic property),
Pressure (piezochromic property),
A solvent (solvatochromic property),
A gas (chimiochromic property),
The expertise of OliKrom allows for each of these properties:
To adjust the request colors,
To obtain reversible and/or irreversible color-shifting,
To modulate the speed of the color change,
To control the issues of fatigability.
To insert these adaptive pigments in a formulation (paint, ink, masterbatch, …) without altering the properties!
To produce on an industrial scale paintings, inks, master batches, …
Applications
SAFETY
Threshold temperature indicators / industrial pipes, thermal mapping.
Control: Temperature Indicator for monitoring sensitive products: cold chain, transport & medical vaccines or blood products.
Sterilization indicator: labels or inks.
Adhesives: indicator of adhesion, optimum drying.
Food Packaging: temperature indicator for the consumption of a product: beer, wine, vodka, champagne, cans and bottles, hot and cold drinks, baby food.
TRACEABILITY / INFRINGEMENT
Irreversible overheat indicator of industrial processes.
Security inks: offset ink for ticketing, games, secure access badges.
Plastic toys: decor with changing color, labels, packaging, paper / plastic promotional, “dynamic” advertising inserts.
Cosmetics: Bottles & Jars of cosmetic or perfume.
Smart Textiles: comfort indicator, clarification of the textile with temperature.
Fluorescent Pigments and Phosphorescent Pigments
Source: PHOTOLUMINESCENTS: FLUORESCENT AND PHOSPHORESCENT INKS AND PAINTS / OliKrom
Photochromic Pigments
Piezochromic Pigments
Thermochromic
Type
Reversible Thermochromic Material
Irreversible Thermochromic Material
Material
Liquid Crystal
Leuco Dyes
Pigment
Other Materials
Application
Roof Coatings
Printing
Food Packaging
Cosmetics
Other Applications
Solvachromes and Chemochromes
Color Change in Biology
Animals
Chameleon
Golden Tortoise Beetle
Mimic Octopus
Pacific Tree Frog
Sea Horses
Flounders
Cuttlefish
Crab Spiders
Squid
Cyanea Octopus
Mechanisms for Color Change
Chromatophores
Leucophores
Iridophores
Source: Adaptive camouflage helps blend into the environment
Cephalopods such as cuttlefish often use use adaptive camouflage to blend in with their surroundings. They are able to match colors and surface textures of their surrounding environments by adjusting the pigment and iridescence of their skin.
On the skin surface, chromatophores (tiny sacs filled with red, yellow, or brown pigment) absorb light of various wavelengths. Once visual input is processed, the cephalopod sends a signal to a nerve fiber, which is connected to a muscle. That muscle relaxes and contracts to change the size and shape of the chromatophore. Each color chromatophore is controlled by a different nerve, and when the attached muscle contracts, it flattens and stretches the pigment sack outward, expanding the color on the skin. When that muscle relaxes, the chromatophore closes back up, and the color disappears. As many as two hundred of these may fill a patch of skin the size of a pencil eraser, like a shimmering pixel display.
The innermost layer of skin, composed of leucophores, reflects ambient light. These broadband light reflectors give the cephalopods a ‘base coat’ that helps them match their surroundings.
Between the colorful chromatophores and the light-scattering leucophores is a reflective layer of skin made up of iridophores. These reflect light to create pink, yellow, green, blue, or silver coloration, while the reflector cells (found only in octopuses) reflect blue or green.
The mimic octopus changes their skin tone and body shape to copy other sea creatures.
There are a few animals that have the unique ability to change colors. The ability to change colors can help animals protect themselves against their predators because it allows them to blend into their natural environment. Here is a list of 10 color changing animals.
10. Chameleon
A chameleon is a unique species of lizard famous for changing its skin color. It does so to camouflage with its surrounding. Sometimes chameleons change their color when they are angry or fearful. To change its color, the chameleon adjusts a layer of specialized cells underlying its skin. Others change color in response to humidity, light, and temperature. Chameleons never stop growing. They keep shedding their skin from time to time. Furthermore, chameleons have excellent eyesight characterized by a 360-degree arc vision. Although chameleons do not hear, their bodies detect sound within the surrounding.
9. Golden Tortoise Beetle
The golden tortoise beetle is an insect that can change its color. The species with this ability include Charidotella sexpunctata and Charidotella egregia. The tortoise beetles change color due to particular events that occur in their environment. Such events include meeting a willing mate and being touched by a curious human being. Hence, when they are mating or agitated, the tortoise beetles change their color from gold to a bright red color. The change of color occurs due to a process referred to as optical illusion.
8. Mimic Octopus
Mimic octopus, scientifically known as Thaumoctopus mimicus, change their color and they can also mimic other sea creatures such as a lionfish, jellyfish, stingrays, and sea snakes. The mimic octopus can pick the color of the sea creature that they intend to mimic. The mimic octopuses change their body shape to avoid potential predators. The change of skin color helps them to adapt to their surrounding. Mimic octopuses can change color and mimic shapes due to their skin which is very responsive to the environment.
7. Pacific Tree Frog
The Pacific Tree Frog inhabits North America. One of its common features is the sticky toe pads. The sticky toe pads enable them to climb trees and plants. The Pacific Tree Frog changes its color to blend in with its surroundings. The change of color is a defense mechanism against predators such as raccoons, bullfrogs, snakes, heron, and many others. Pacific Tree Frogs also change their color based on the seasons and temperature. When the temperatures are high, they turn into a shade of yellow. An example of Pacific Tree Frog species that changes color is Hyla regilla. The process of color change in Pacific Tree Frogs takes 1-2 minutes.
6. Seahorses
Seahorses, such as the thorny seahorse, are among the marine animals that have mastered changing their color. The purpose of changing their skin color is to camouflage, frighten predators, communicate their emotions, and for courtship. Complex interactions between the brain, nervous system, hormones, and organelles make it possible for the seahorses to change their color. The organelles responsible for these color changes are known as chromatophores. Regarding the speed at which the skin color changes, this depends on the stimulus. For instance, in a life or death situation such as involving a predator, the color changes quickly. But whenever the seahorse is courting a mate, the change takes place slowly.
5. Flounders
Flounders are naturally brown. However, they can change color to suit their surroundings. A flounder uses its vision and specialized cells inside the skin to change color. The cells, in turn, have color pigments and are linked to the eyes of the flounders. When a flounder moves to a new environment, the retina in the eyes captures the new color. Consequently, the color seen by the eyes are transmitted to the cells. The cells adjust the pigmentation to match the surface color. Scientists have discovered that flounders depend entirely on their vision to change color. When their eyes are damaged, then they have difficulties in camouflaging to their surrounding. An example of flounder species that changes color is the peacock flounder.
4. Cuttlefish
Cuttlefish are cephalopods that change color to feed on prey and avoid predators craftily. They have three mechanisms by which they can change color. Firstly, the cuttlefish skin contains papillae that alter the tone of the fish. The papillae cause the skin to become smooth or rough depending on the environment. Secondly, camouflaging is possible because of the chromatophores in their skin. The chromatophores are sacs of color pigments. To change color, these sacs receive color-changing instructions from the brain and act accordingly. Lastly, cuttlefish have reflecting plates called leucophores and iridophores. The plates enable the fish to change its color.
3. Crab Spiders
Spiders called flower spiders (or crab spiders) change their color. They usually change color to hide from their prey. Consequently, the spiders change color to resemble the flower surface on which they sit through the reflection of light. Some spiders release a yellow pigment that enhances their color changing process. An example of a species of spider with such color changing features is Misumenoides formosipes and Misumena vatia. The color change from white to yellow takes 10-25 days. Hence, the flower spiders patiently wait for the completion of the process before they can attack their prey.
2. Squid
Squids are marine cephalopods. They possess two long tentacles and eight arms. An interesting fact about the squids is that their blood is blue. Furthermore, they have three hearts instead of one like other fish. The squids are uniquely beautiful and able to change color. They change color using chromatophores engraved in their skin. The purpose of changing color is to match the surface they are on so that they can avoid predators. The camouflage also acts as a hunting tactic since it enables them to hide away from their prey.
1. Cyanea Octopus
Known as the big blue octopus or the day octopus, octopus cyabea is found in the waters of the Indo-Pacific. It is known as the day octopus as it is most active during the daytime in contrast to most other octopus species. The cyanea octopusis especially adept at camouflage, able to not only frequently change the color of their skin, but also recreate patterns and textures. On the hunt for crabs, molluscs, shrimp, and fish, the cyanea octopus is able to quickly adapt its appearance to its surroundings, even mimicking moving shadows such as overhead clouds.
Color Change in Plants And Flowers
Color change in Leaves and Flowers
Chlorophyll – Green
Cartenoids – Xanthophylls – Yellow as in Corn
Cartenoids – Carotenes – Orange as in Carrots
Anthrocyanins – Blueberries and Cherries – Blue, purple, red, pink
Flavonols – Pale yellows and whites
Plants change colors
Change in Heat
Change in pH
During the Fall
During the day
Color Fading and Color Metamerism are also important problems but are not discussed in this post.
Source: The science behind why leaves change color in autumn
A rainbow of autumn colors
The green color of chlorophyll is so strong that it masks any other pigment. The absence of green in the fall lets the other colors come through. Leaves also contain the pigments called carotenoids; xanthophylls are yellow (such as in corn) and carotenes are orange (like in carrots). Anthocyanins (also found in blueberries, cherries) are pigments that are only produced in the fall when it is bright and cold. Because the trees cut off most contact with their leaves at this point, the trapped sugar in the leaves’ veins promotes the formation of anthocyanins, which are used for plant defense and create reddish colors.
However, trees in the fall aren’t just yellow and red: they are brown, golden bronze, golden yellow, purple-red, light tan, crimson, and orange-red. Different trees have different proportions of these pigments; the amount of chlorophyll left and the proportions of other pigments determine a leaf’s color. A combination of anthocyanin and chlorophyll makes a brown color, while anthocyanins plus carotenoids create orange leaves.
Source: The science behind why leaves change color in autumn
Can Hibiscus Change Color: Reasons For Hibiscus Turning A Different Color
07/20/20
Can hibiscus change color? The Confederate Rose (Hibiscus mutabilis) is famous for its dramatic color changes, with flowers that can go from white to pink to deep red within one day. But almost all hibiscus varieties produce flowers that can change colors under certain circumstances. Read on to learn more.
Reasons for Color Changing in Hibiscus
If you’ve ever noticed the flowers on your hibiscus turning a different color, you’ve probably wondered what was behind the change. To understand why this happens, we need to look at what creates flower colors in the first place.
Three groups of pigments create the vibrant color displays of hibiscus flowers. Anthocyanins produce blue, purple, red, and pink colors, depending on the individual pigment molecule and the pH it is exposed to. Flavonols are responsible for pale yellow or white colors. Carotenoids create colors on the “warm” side of the spectrum – yellows, oranges, and reds.
Each hibiscus variety has its own genetics that determine what pigments, and what range of colors it can produce. However, within that range, temperature, sunlight, pH, and nutrition can all affect the levels of different pigments in a flower and what color they appear.
The blue- and red-colored anthocyanins are water-soluble pigments carried in plant sap. Meanwhile, the red, orange and yellow carotenoids are fat-soluble pigments created and stored in the plastids (compartments in plant cells similar to the chloroplasts that carry out photosynthesis). Therefore, anthocyanins are less protected and more sensitive to environmental changes, while carotenoids are more stable. This difference helps explain the color changes in hibiscus.
Anthocyanins exposed to hot conditions will often break down, causing flower colors to fade, while carotenoid-based colors hold up well in the heat. High temperatures and bright sunlight also enhance carotenoid production, leading to bright reds and oranges.
On the other hand, plants produce more anthocyanins in cold weather, and the anthocyanins they produce tend to be more red- and pink-colored as opposed to blue or purple. For this reason, some anthocyanin dependent hibiscus flowers will produce brilliant color displays during cool weather or in partial shade, but will fade in bright, hot sunlight.
Similarly, flavonols exposed to high temperatures will fade from yellow to white, while cold weather will cause an increase in production and a deepening of yellow flower colors.
Other Factors in Hibiscus Color Change
Some anthocyanin pigments will change color depending on the pH they’re exposed to within the flower. The pH doesn’t usually change over time within a hibiscus flower because it is determined genetically, but patches of different pH levels can lead to multiple colors occurring within one flower.
Nutrition is also a factor in color changes. Adequate sugar and protein in the sap are required for anthocyanin production. Making sure your plant has enough fertility and nutrients is important for vibrant colors in anthocyanin dependent flowers.
So, depending on its variety, your hibiscus changed color because of some combination of temperature, sunlight, nutrition, or pH has taken place. Can gardeners control this hibiscus color change? Yes, indirectly – by controlling the plant’s environment: shade or sun, good fertility, and protection from hot or cold weather.
Anthocyanins and carotenoids… plus some other things.
Flowers come in all shapes and sizes, but what makes them truly stand apart from each other is their vibrant colors. These colors are made up of pigments and, generally speaking, the fewer the pigments, the lighter the color. The most common pigments in flowers come in the form of anthocyanins. These pigments range in color from white to red to blue to yellow to purple and even black and brown. A different kind of pigment class is made up of the carotenoids. Carotenoids are responsible for some yellows, oranges, and reds. (These little guys are what cause the brilliant colors of autumn leaves!) While many flowers get their colors from either anthocyanins or carotenoids, there are some that can get their colors from a combination of both.
Anthocyanins and carotenoids are the main sources of flower coloration, but there are other factors that can affect how colors present themselves. The amount of light flowers receive while they grow, the temperature of the environment around them, even the pH level of the soil in which they grow can affect their coloration. Another factor is stress from the environment. This stress can include a drought or a flood or even a lack of nutrition in the soil, all of which can dampen the coloration of flowers. And then, of course, there is the visual that the eye and brain form together: humans can, for the most part, view all colors in the visible spectrum, BUT every human perceives color differently, so a red rose may appear more vibrant to one person while it appears more muted to another. Beauty (and color!) is in the eye of the beholder.
Silvia Vignolini1,2, Edwige Moyroud3, Beverley J. Glover3 and Ullrich Steiner1
1Cavendish Laboratory, University of Cambridge, JJ Thomson Avenue, Cambridge CB3 0HE, UK 2Department of Physics and Astronomy, University College London, Gower Street, London WC1E 6BT, UK 3Department of Plant Sciences, University of Cambridge, Downing Street, Cambridge CB2 3EA, UK
What happens in the Present is not only determined by the Past but also by the Future. Karma and Destiny both play a role as to what is going on in your life Now.
Key Terms
Recursion
Incursion
Hyper Incursion
Discrete Processes
Cellular Automata
Fractal Machine
Hypersets
Interpenetration
Turing Machine
Symmetry
Non Well Founded Set Theory
Sets as Graphs
Leela
Predetermined Future
Bhagya
Fate
Destiny
Karma
Anticipation
Four Causes of Aristotle
Material Cause
Formal Cause
Efficient Cause
Final Cause
Left Computer
Right Computer
Parallel Computing
Fifth and the Fourth in Music Theory
Bicameral Brain
Hemispheric Division of Brain
One, Two, Three. Where is the Fourth?
From GENERATION OF FRACTALS FROM INCURSIVE AUTOMATA, DIGITAL DIFFUSION AND WAVE EQUATION SYSTEMS
The recursion consists of the computation of the future value of the variable vector X(t+l) at time t+l from the values of these variables at present and/or past times, t, t-l, t-2 ….by a recursive function :
X (t+ 1) =f(X(t), X(t-1) …p..)
where p is a command parameter vector. So, the past always determines the future, the present being the separation line between the past and the future.
Starting from cellular automata, the concept of Fractal Machines was proposed in which composition rules were propagated along paths in the machine frame. The computation is based on what I called “INclusive reCURSION”, i.e. INCURSION (Dubois, 1992a- b). An incursive relation is defined by:
X(t+l) =f(…, X (t+l), X(t), X(t-1) ..p..).
which consists in the computation of the values of the vector X(t+l) at time t+l from the values X(t-i) at time t-i, i=1, 2 …. , the value X(t) at time t and the value X(t+j) at time t+j, j=l, 2, …. in function of a command vector p. This incursive relation is not trivial because future values of the variable vector at time steps t+l, t+2 …. must be known to compute them at the time step t+ 1.
In a similar way to that in which we define hyper recursion when each recursive step generates multiple solutions, I define HYPERINCURSION. Recursive computational transformations of such incursive relations are given in Dubois and Resconi (1992, 1993a-b).
I have decided to do this for three reasons. First, in relativity theory space and time are considered as a four-vector where time plays a role similar to space. If time t is replaced by space s in the above definition of incursion, we obtain
X(s+ l) =f( …, X(s+ 1), X(s), X (s-l) …p.).
and nobody is astonished: a Laplacean operator looks like this. Second, in control theory, the engineers control engineering systems by defining goals in the future to compute their present state, similarly to our haman anticipative behaviour (Dubois, 1996a-b). Third, I wanted to try to do a generalisation of the recursive and sequential Turing Machine in looking at space-time cellular automata where the order in which the computations are made is taken into account with an inclusive recursion.
We have already proposed some methods to realise the design of any discrete systems with an extension of the recursion by the concept of incursion and hyperincursion based on the Fractal Machine, a new type of Cellular Automata, where time plays a central role. In this framework, the design of the model of any discrete system is based on incursion relations where past, present and future states variables are mixed in such a way that they define an indivisible wholeness invariant. Most incursive relations can be transformed in different sets of recursive algorithms for computation. In the same way, the hyperincursion is an extension of the hyper recursion in which several different solutions can be generated at each time step. By the hyperincursion, the Fractal Machine could compute beyond the theoretical limits of the Turing Machine (Dubois and Resconi, 1993a-b). Holistic properties of the hyperincursion are related to the Golden Ratio with the Fibonacci Series and the Fractal Golden Matrix (Dubois and Resconi, 1992). An incursive method was developed for the inverse problem, the Newton- Raphson method and an application in robotics (Dubois and Resconi, 1995). Control by incursion was applied to feedback systems (Dubois and Resconi, 1994). Chaotic recursions can be synchronised by incursion (1993b). An incursive control of linear, non- linear and chaotic systems was proposed (Dubois, 1995a, Dubois and Resconi, 1994, 1995). The hyperincursive discrete Lotka-Voiterra equations have orbital stability and show the emergence of chaos (Dubois, 1992). By linearisation of this non-linear system, hyperincursive discrete harmonic oscillator equations give stable oscillations and discrete solutions (Dubois, 1995). A general theory of stability by incursion of discrete equations systems was developed with applications to the control of the numerical instabilities of the difference equations of the Lotka-Volterra differential equations as well as the control of the fractal chaos in the Pearl-Verhulst equation (Dubois and Resconi, 1995). The incursion harmonic oscillator shows eigenvalues and wave packet like in quantum mechanics. Backward and forward velocities are defined in this incursion harmonic oscillator. A connection is made between incursion and relativity as well as the electromagnetic field. The foundation of a hyperincursive discrete mechanics was proposed in relation to the quantum mechanics (Dubois and Resconi, 1993b, 1995).
This paper will present new developments and will show that the incursion and hyper-incursion could be a new tool of research and development for describing systems where the present state of such systems is also a function of their future states. The anticipatory property of incursion is an incremental final cause which could be related to the Aristotelian Final Cause.
Aristotle identified four explicit categories of causation: 1. Material cause; 2. Formal cause; 3. Efficient cause; 4. Final cause. Classically, it is considered that modem physics and mechanics only deal with efficient cause and biology with material cause. Robert Rosen (1986) gives another interpretation and asks why a certain Newtonian mechanical system is in the state (phase) Ix(t) (position), v(t) (velocity)]:
1. Aristotle’s “material cause” corresponds to the initial conditions of the system [x(0), v(0)] at time t=0.
2. The current cause at the present time is the set of constraints which convey to the system an “identity”, allowing it to go by recursion from the given initial phase to the latter phase, which corresponds to what Aristotle called formal cause.
3. What we call inputs or boundary conditions are the impressed forces by the environment, called efficient cause by Aristotle.
As pointed out by Robert Rosen, the first three of Aristotle’s causal categories are tacit in the Newtonian formalism: “the introduction of a notion of final cause into the Newtonian picture would amount to allowing a future state or future environment to affect change of state in the present, and this would be incompatible with the whole Newtonian picture. This is one of the main reasons that the concept of Aristotelian finality is considered incompatible with modern science.
In modern physics, Aristotelian ideas of causality are confused with determinism, which is quite different…. That is, determinism is merely a mathematical statement of functional dependence or linkage. As Russell points out, such mathematical relations, in themselves, carry no hint as to which of their variables are dependent and which are independent.”
The final cause could impress the present state of evolving systems, which seems a key phenomenon in biological systems so that the classical mathematical models are unable to explain many of these biological systems. An interesting analysis of the Final Causation was made by Emst von Glasersfeld (1990). The self-referential fractal machine shows that the hyperincursive field dealing with the final cause could be also very important in physical and computational systems. The concepts of incursion and hyper-incursion deal with an extension of the recursive processes for which future states can determine present states of evolving systems. Incursion is defined as invariant functional relations from which several recursive models with interacting variables can be constructed in terms of diverse physical structures (Dubois & Resconi, 1992, 1993b). Anticipation, viewed as an Aristotelian final cause, is of great importance to explain the dynamics of systems and the semantic information (Dubois, 1996a-b). Information is related to the meaning of data. It is important to note that what is usually called Information Theory is only a communication theory dealing with the communication of coded data in channels between a sender and a receptor without any reference to the semantic aspect of the messages. The meaning of the message can only be understood by the receiver if he has the same cultural reference as the sender of the message and even in this case, nobody can be sure that the receiver understands the message exactly as the sender. Because the message is only a sequential explanation of a non-communicable meaning of an idea in the mind of the sender which can be communicated to the receiver so that a certain meaning emerges in his mind. The meaning is relative or subjective in the sense that it depends on the experiential life or imagination of each of us. It is well- known that the semantic information of signs (like the coding of the signals for traffic) are the same for everybody (like having to stop at the red light at a cross roads) due to a collective agreement of their meaning in relation to actions. But the semantic information of an idea, for example, is more difficult to codify. This is perhaps the origin of creativity for which a meaning of something new emerges from a trial to find a meaning for something which has no a priori meaning or a void meaning.
Mind dynamics seems to be a parallel process and the way we express ideas by language is sequential. Is the sequential information the same as the parallel information? Let us explain this by considering the atoms or molecules in a liquid. We can calculate the average velocity of the particles from in two ways. The first way is to consider one particular particle and to measure its velocity during a certain time. One obtains its mean velocity which corresponds to the mean velocity of any particle of the liquid. The sec- ond way is to consider a certain number of particles at a given time and to measure the velocity of each of them. This mean velocity is equal to the first mean velocity. So there are two ways to obtain the same information. One by looking at one particular element along the time dimension and the other by looking at many elements at the same time. For me, explanation corresponds to the sequential measure and understanding to the parallel measure. Notice that ergodicity is only available with simple physical systems, so in general we can say that there are distortions between the sequential and the parallel view of any phenomenon. Perhaps the brain processes are based on ergodicity: the left hemisphere works in a sequential mode while the right hemisphere works in a parallel mode. The left brain explains while the right brain understands. The two brains arecomplementary and necessary.
Today computer science deals with the “left computer”. Fortunately, the informaticians have invented parallel computers which are based on complex multiplication of Turing Machines. It is now the time to reconsider the problem of looking at the “right computer”. Perhaps it will be an extension of the Fractal Machine (Dubois & Resconi, 1993a).
I think that the sequential way deals with the causality principle while the parallel way deals with a finality principle. There is a paradox: causality is related to the successive events in time while finality is related to a collection of events at a simultaneous time, i.e. out of time.Causality is related to recursive computations which give rise to the local generation of patterns in a synchronic way. Finality is related to incursive or hyperincursive symmetry invariance which gives rise to an indivisible wholeness, a holistic property in a diachronic way. Recursion (and Hyper recursion) is defined in the Sets Theory and Incursion (and Hyperincursion) could be defined in the new framework of the Hypersets Theory (Aczel, 1987; Barwise, Moss, 1991).
If the causality principle is rather well acknowledged, a finality principle is still controversial. It would be interesting to re-define these principles. Causality is defined for sequential events. If x(t) represents a variable at time t, a causal rule x(t+l) = f(x(t)) gives the successive states of the variable x at the successive time steps t, t+l, t+2, … from the recursive functionf(x(t)), starting with an initial state x(0) at time t=0. Defined like this, the system has no degrees of freedom: it is completely determined by the function and the initial condition. No new things can happen for such a system: the whole future is completely determined by its past. It is not an evolutionary system but a developmental system. If the system tends to a stable point, x(t+l) = x(t) and it remains in this state for ever. The variable x can represent a vector of states as a generalisation.
In the same way, I think that determinism is confused with predictability, in modern physics. The recent fractal and deterministic chaos theory (Mandeibrot, 1982; Peitgen, Jurgens, Saupe, 1992) is a step beyond classical concepts in physics. If the function is non-linear, chaotic behaviour can appear, what is called (deterministic) chaos. In this case, determinism does not give an accurate prediction of the future of the system from its initial conditions, what is called sensitivity to initial conditions. A chaotic system loses the memory of its past by finite computation. But it is important to point out that an average value, or bounds within which the variable can take its values, can be known;
it is only the precise values at the successive steps which are not predictable. The local information is unpredictable while the global symmetry is predictable. Chaos can presents a fractai geometry which shows a self-similarity of patterns at any scale.
A well-known fractal is the Sierpinski napkin. The self-similarity of pattems at any scale can be viewed as a symmetry invariance at any scale. An interesting property of such fractals is the fact that the final global pattern symmetry can be completely independent of the local pattern symmetry given as the initial condition of the process from which the fractal is built. The symmetry of the fractal structure, a final cause, can be independent of the initial conditions, a material cause. The formal cause is the local symmetry of the generator of the fractal, independently of its material elements and the efficient cause can be related to the recursive process to generate the fractal. In this particular fractal geometry, the final cause is identical to the final cause. The efficient cause is the making of the fractal and the material cause is just a substrate from which the fractal emerges but this substrate doesn’t play a role in the making.
Finally, the concepts of incursion and hyperincursion can be related to the theory of hypersets which are defined as sets containing themselves. This theory of hypersets is an alternative theory to the classical set theory which presents some problems as the in- completeness of G6del: a formal system cannot explain all about itself and some propositions cannot be demonstrated as true or false (undecidability). Fundamental entities of systems which are considered as ontological could be explain in a non-ontological way by self-referential systems.
In mathematics, a knot is defined as a closed, non-self-intersecting curve that is embedded in three dimensions and cannot be untangled to produce a simple loop (i.e., the unknot). While in common usage, knots can be tied in string and rope such that one or more strands are left open on either side of the knot, the mathematical theory of knots terms an object of this type a “braid” rather than a knot. To a mathematician, an object is a knot only if its free ends are attached in some way so that the resulting structure consists of a single looped strand.
A knot can be generalized to a link, which is simply a knotted collection of one or more closed strands.
The study of knots and their properties is known as knot theory. Knot theorywas given its first impetus when Lord Kelvin proposed a theory that atoms were vortex loops, with different chemical elements consisting of different knotted configurations (Thompson 1867). P. G. Tait then cataloged possible knots by trial and error. Much progress has been made in the intervening years.
Schubert (1949) showed that every knot can be uniquely decomposed (up to the order in which the decomposition is performed) as a knot sum of a class of knots known as prime knots, which cannot themselves be further decomposed (Livingston 1993, p. 5; Adams 1994, pp. 8-9). Knots that can be so decomposed are then known as composite knots. The total number (prime plus composite) of distinct knots (treating mirror images as equivalent) having , 1, … crossings are 1, 0, 0, 1, 1, 2, 5, 8, 25, … (OEIS A086825).
Klein proved that knots cannot exist in an even-dimensional space . It has since been shown that a knot cannot exist in any dimension . Two distinct knots cannot have the same knot complement (Gordon and Luecke 1989), but two links can! (Adams 1994, p. 261).
Knots are most commonly cataloged based on the minimum number of crossings present (the so-called link crossing number). Thistlethwaite has used Dowker notation to enumerate the number of prime knots of up to 13 crossings, and alternating knots up to 14 crossings. In this compilation, mirror images are counted as a single knot type. Hoste et al. (1998) subsequently tabulated all prime knots up to 16 crossings. Hoste and Weeks subsequently began compiling a list of 17-crossing prime knots (Hoste et al. 1998).
Another possible representation for knots uses the braid group. A knot with crossings is a member of the braid group.
There is no general algorithm to determine if a tangled curve is a knot or if two given knots are interlocked. Haken (1961) and Hemion (1979) have given algorithms for rigorously determining if two knots are equivalent, but they are too complex to apply even in simple cases (Hoste et al. 1998).
LH Kauffman with Trefoil Knot in the back.
From Reflexivity
A Knot
Trefoil Knot
From Reflexivity
This slide show has been only an introduction to certain mathematical and conceptual points of view about reflexivity.
In the worlds of scientific, political and economic action these principles come into play in the way structures rise and fall in the play of realities that are created from (almost) nothing by the participants in their desire to profit, have power or even just to have clarity and understanding. Beneath the remarkable and unpredictable structures that arise from such interplay is a lambent simplicity to which we may return, as to the source of the world.
From Laws of Form and the Logic of Non-Duality
This talk will trace how a mathematics of distinction arises directly from the process of discrimination and how that language, understood rightly as an opportunity to join as well as to divide, can aid in the movement between duality and non-duality that is our heritage as human beings on this planet.The purpose of this talk is to express this language and invite your participation in it and to present the possiblity that all our resources physical, scientific, logical, intellectual, empathic are our allies in the journey to transcend separation.
Codes and conventions are the basis of our social life and from time immemorial have divided the world of culture from the world of nature. The rules of grammar, the laws of government, the precepts of religion, the value of money, the rules of chess etc., are all human conventions that are profoundly different from the laws of physics and chemistry, and this has led to the conclusion that there is an unbridgeable gap between nature and culture. Nature is governed by objective immutable laws, whereas culture is produced by the mutable conventions of the human mind.
In this millennia-old framework, the discovery of the genetic code, in the early 1960s, came as a bolt from the blue, but strangely enough it did not bring down the barrier between nature and culture. On the contrary, a protective belt was quickly built around the old divide with an argument that effectively emptied the discovery of all its revolutionary potential. The argument that the genetic code is not a real code because its rules are the result of chemical affinities between codons and amino acids and are therefore determined by chemistry. This is the ‘Stereochemical theory’, an idea first proposed by George Gamow in 1954, and re-proposed ever since in many different forms (Pelc and Welton 1966; Dunnil 1966; Melcher 1974; Shimizu 1982; Yarus 1988, 1998; Yarus, Caporaso and Knight 2005). More than fifty years of research have not produced any evidence in favour of this theory and yet the idea is still circulating, apparently because of the possibility that stereochemical interactions might have been important at some early stages of evolution (Koonin and Novozhilov 2009). The deep reason is probably the persistent belief that the genetic code must have been a product of chemistry and cannot possibly be a real code. But what is a real code?
The starting point is the idea that a code is a set of rules that establish a correspondence, or a mapping, between the objects of two independent worlds (Barbieri 2003). The Morse code, for example, is a mapping between the letters of the alphabet and groups of dots and dashes. The highway code is a correspondence between street signals and driving behaviours (a red light means ‘stop’, a green light means ‘go’, and so on).
What is essential in all codes is that the coding rules, although completely compatible with the laws of physics and chemistry, are not dictated by these laws. In this sense they are arbitrary, and the number of arbitrary relationships between two independent worlds is potentially unlimited. In the Morse code, for example, any letter of the alphabet could be associated with countless combinations of dots and dashes, which means that a specific link between them can be realized only by selecting a small number of rules. And this is precisely what a code is: a small set of arbitrary rules selected from a potentially unlimited number in order to ensure a specific correspondence between two independent worlds.
This definition allows us to make experimental tests because organic codes are relationships between two worlds of organic molecules and are necessarily implemented by a third type of molecules, called adaptors, that build a bridge between them. The adaptors are required because there is no necessary link between the two worlds, and a fixed set of adaptors is required in order to guarantee the specificity of the correspondence. The adaptors, in short, are the molecular fingerprints of the codes, and their presence in a biological process is a sure sign that that process is based on a code.
This gives us an objective criterion for discovering organic codes and their existence is no longer a matter of speculation. It is, first and foremost, an experimental problem. More precisely, we can prove that an organic code exists, if we find three things: (1) two independents worlds of molecules, (2) a set of adaptors that create a mapping between them, and (3) the demonstration that the mapping is arbitrary because its rules can be changed, at least in principle, in countless different ways.
Two outstanding examples
The genetic code
In protein synthesis, a sequence of nucleotides is translated into a sequence of amino acids, and the bridge between them is realized by a third type of molecules, called transfer-RNAs, that act as adaptors and perform two distinct operations: at one site they recognize groups of three nucleotides, called codons, and at another site they receive amino acids from enzymes called aminoacyl-tRNA-synthetases. The key point is that there is no deterministic link between codons and amino acids since it has been shown that any codon can be associated with any amino acid (Schimmel 1987; Schimmel et al. 1993). Hou and Schimmel (1988), for example, introduced two extra nucleotides in a tRNA and found that that the resulting tRNA was carrying a different amino acid. This proved that the number of possible connections between codons and amino acids is potentially unlimited, and only the selection of a small set of adaptors can ensure a specific mapping. This is the genetic code: a fixed set of rules between nucleic acids and amino acids that are implemented by adaptors. In protein synthesis, in conclusion, we find all the three essential components of a code: (1) two independents worlds of molecules (nucleotides and amino acids), (2) a set of adaptors that create a mapping between them, and (3) the proof that the mapping is arbitrary because its rules can be changed.
The signal transduction codes
Signal transduction is the process by which cells transform the signals from the environment, called first messengers, into internal signals, called second messengers. First and second messengers belong to two independent worlds because there are literally hundreds of first messengers (hormones, growth factors, neurotransmitters, etc.) but only four great families of second messengers (cyclic AMP, calcium ions, diacylglycerol and inositol trisphosphate) (Alberts et al. 2007). The crucial point is that the molecules that perform signal transduction are true adaptors. They consists of three subunits: a receptor for the first messengers, an amplifier for the second messengers, and a mediator in between (Berridge 1985). This allows the transduction complex to perform two independent recognition processes, one for the first messenger and the other for the second messenger. Laboratory experiments have proved that any first messenger can be associated with any second messenger, which means that there is a potentially unlimited number of arbitrary connections between them. In signal transduction, in short, we find all the three essential components of a code: (1) two independents worlds of molecules (first messengers and second messengers), (2) a set of adaptors that create a mapping between them, and (3) the proof that the mapping is arbitrary because its rules can be changed (Barbieri 2003).
A world of organic codes
In addition to the genetic code and the signal transduction codes, a wide variety of new organic codes have come to light in recent years. Among them: the sequence codes (Trifonov 1987, 1989, 1999), the Hox code (Paul Hunt et al. 1991; Kessel and Gruss 1991), the adhesive code (Redies and Takeichi 1996; Shapiro and Colman 1999), the splicing codes (Barbieri 2003; Fu 2004; Matlin et al. 2005; Pertea et al. 2007; Wang and Burge 2008; Barash et al. 2010; Dhir et al. 2010), the signal transduction codes (Barbieri 2003), the histone code (Strahl and Allis 2000; Jenuwein and Allis 2001; Turner 2000, 2002, 2007; Kühn and Hofmeyr 2014), the sugar code (Gabius 2000, 2009), the compartment codes (Barbieri 2003), the cytoskeleton codes (Barbieri 2003; Gimona 2008), the transcriptional code (Jessell 2000; Marquard and Pfaff 2001; Ruiz i Altaba et al. 2003; Flames et al. 2007), the neural code (Nicolelis and Ribeiro 2006; Nicolelis 2011), a neural code for taste (Di Lorenzo 2000; Hallock and Di Lorenzo 2006), an odorant receptor code(Dudai 1999; Ray et al. 2006), a space code in the hippocampus (O’Keefe and Burgess 1996, 2005; Hafting et al. 2005; Brandon and Hasselmo 2009; Papoutsi et al. 2009), the apoptosis code (Basañez and Hardwick 2008; Füllgrabe et al. 2010), the tubulin code (Verhey and Gaertig 2007), the nuclear signalling code (Maraldi 2008), the injective organic codes (De Beule et al. 2011), the molecular codes (Görlich et al. 2011; Görlich and Dittrich 2013), the ubiquitin code (Komander and Rape 2012), the bioelectric code (Tseng and Levin 2013; Levin 2014), the acoustic codes (Farina and Pieretti 2014), the glycomic code (Buckeridge and De Souza 2014; Tavares and Buckeridge 2015) and the Redox code (Jones and Sies 2015).
The living world, in short, is literally teeming with organic codes, and yet so far their discoveries have only circulated in small circles and have not attracted the attention of the scientific community at large.
Code Biology
Code Biology is the study of all codes of life with the standard methods of science. The genetic code and the codes of culture have been known for a long time and represent the historical foundation of Code Biology. What is really new in this field is the study of all codes that came after the genetic code and before the codes of culture. The existence of these codes is an experimental fact – let us never forget this – but also more than that. It is one of those facts that have extraordinary theoretical implications.
The first is the role that the organic codes had in the history of life. The genetic code was a precondition for the origin of the first cells, the signal transduction codes divided the descendants of the common ancestor into the primary kingdoms of Archaea, Bacteria and Eukarya, the splicing codes were instrumental to the origin of the nucleus, the histone code provided the rules of chromatin, and the cytoskeleton codes allowed the Eukarya to perform internal movements, including those of mitosis and meiosis (Barbieri 2003, 2015). The greatest events of macroevolution, in other words, were associated with the appearance of new organic codes, and this gives us a completely new understanding of the history of life.
The second great implication is the fact that the organic codes have been highly conserved in evolution, which means that they are the great invariants of life, the sole entities that have been perpetuated while everything else has been changed. Code Biology, in short, is uncovering a new history of life and bringing to light new fundamental concepts. It truly is a new science, the exploration of a vast and still largely unexplored dimension of the living world, the real new frontier of biology.
References
Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2007) Molecular Biology of the Cell. 5th Ed. Garland, New York.
Barash Y, Calarco JA, Gao W, Pan Q, Wang X, Shai O, Blencow BJ and Frey BJ (2010). Deciphering the splicing code. Nature, Vol 465, 53-59.
Barbieri M (2003) The Organic Codes. An Introduction to Semantic Biology. Cambridge University Press, Cambridge, UK.
Barbieri M (2015) Code Biology. A New Science of Life. Springer, Dordrecht.
Basañez G and Hardwick JM (2008) Unravelling the Bcl-2 Apoptosis Code with a Simple Model System. PLoS Biol 6(6): e154. Doi: 10.137/journal.pbio.0060154.
Berridge M (1985) The molecular basis of communication within the cell. Scientific American, 253, 142-152.
Brandon MP and Hasselmo ME (2009) Sources of the spatial code within the hippocampus. Biology Reports, 1, 3-7.
Buckeridge MS and De Souza AP (2014) Breaking the “Glycomic Code” of cell wall polysaccharides may improve second-generation bioenergy production from biomass. BioEnergy Research, 7, 1065-1073.
De Beule J, Hovig E and Benson M (2011) Introducing Dynamics into the Field of Biosemiotics. Biosemiotics, 4(1), 5-24.
Dhir A, Buratti E, van Santen MA, Lührmann R and Baralle FE, (2010). The intronic splicing code: multiple factors involved in ATM pseudoexon definition. The EMBO Journal, 29, 749–760.
Di Lorenzo PM (2000) The neural code for taste in the brain stem: Response profiles. Physiology and Behaviour, 69, 87-96.
Dudai Y (1999) The Smell of Representations. Neuron 23: 633-635.
Dunnill P (1966) Triplet nucleotide-amino-acid pairing; a stereochemical basis for the division between protein and non-protein amino-acids. Nature, 210, 1267-1268.
Farina A and Pieretti N (2014) Acoustic Codes in Action in a Soundscape Context. Biosemiotics, 7(2), 321–328.
Flames N, Pla R, Gelman DM, Rubenstein JLR, Puelles L and Marìn O (2007) Delineation of Multiple Subpallial Progenitor Domains by the Combinatorial Expression of Transcriptional Codes. The Journal of Neuroscience, 27, 9682–9695.
Fu XD (2004) Towards a splicing code. Cell, 119, 736–738.
Füllgrabe J, Hajji N and Joseph B (2010) Cracking the death code: apoptosis-related histone modifications. Cell Death and Differentiation, 17, 1238-1243.
Gabius H-J (2000) Biological Information Transfer Beyond the Genetic Code: The Sugar Code. Naturwissenschaften, 87, 108-121.
Gabius H-J (2009) The Sugar Code. Fundamentals of Glycosciences. Wiley-Blackwell.
Gamow G (1954) Possible relation between deoxyribonucleic acid and protein structures. Nature, 173, 318.
Gimona M (2008) Protein linguistics and the modular code of the cytoskeleton. In: Barbieri M (ed) The Codes of Life: The Rules of Macroevolution. Springer, Dordrecht, pp 189-206.
Görlich D, Artmann S, Dittrich P (2011) Cells as semantic systems. Biochim Biophys Acta, 1810 (10), 914-923.
Görlich D and Dittrich P (2013) Molecular codes in biological and chemical reaction networks. PLoS ONE 8(1):e54,694, DOI 10.1371/journal.pone.0054694.
Hafting T, Fyhn M, Molden S, Moser MB, Moser EI (2005) Microstructure of a spatial map in the entorhinal cortex. Nature, 436, 801-806.
Hallock RM and Di Lorenzo PM (2006) Temporal coding in the gustatory system. Neuroscience and Behavioral Reviews, 30, 1145-1160.
Hou Y-M and Schimmel P (1988) A simple structural feature is a major determinant of the identity of a transfer RNA. Nature, 333, 140-145.
Hunt P, Whiting J, Nonchev S, Sham M-H, Marshall H, Graham A, Cook M, Alleman R, Rigby PW and Gulisano M (1991) The branchial Hox code and its implications for gene regulation, patterning of the nervous system and head evolution. Development, 2, 63-77.
Jenuwein T and Allis CD (2001) Translating the histone code. Science, 293, 1074-1080.
Jessell TM (2000) Neuronal Specification in the Spinal Cord: Inductive Signals and Transcriptional Codes. Nature Genetics, 1, 20-29.
Jones DP and Sies H (2015) The Redox Code. Antioxidants and Redox Signaling, 23 (9), 734-746.
Kessel M and Gruss P (1991) Homeotic Tansformation of Murine Vertebrae and Concomitant Alteration of Hox Codes induced by Retinoic Acid. Cell, 67, 89-104.
Komander D and Rape M (2012), The Ubiquitin Code. Annu. Rev. Biochem. 81, 203–29.
Koonin EV and Novozhilov AS (2009) Origin and evolution of the genetic code: the universal enigma. IUBMB Life. 61(2), 99-111.
Kühn S and Hofmeyr J-H S (2014) Is the “Histone Code” an organic code? Biosemiotics, 7(2), 203–222.
Levin M (2014) Endogenous bioelectrical networks store non-genetic patterning information during development and regeneration. Journal of Physiology, 592.11, 2295–2305.
Maraldi NM (2008) A Lipid-based Code in Nuclear Signalling. In: Barbieri M (ed) The Codes of Life: The Rules of Macroevolution. Springer, Dordrecht, pp 207-221.
Marquard T and Pfaff SL (2001) Cracking the Transcriptional Code for Cell Specification in the Neural Tube. Cell, 106, 651–654.
Matlin A, Clark F and Smith C (2005) Understanding alternative splicing: towards a cellular code. Nat. Rev. Mol. Cell Biol., 6, 386-398.
Melcher G (1974) Stereospecificity and the genetic code. J. Mol. Evol., 3, 121-141.
Nicolelis M (2011) Beyond Boundaries: The New Neuroscience of Connecting Brains with Machines and How It Will Change Our Lives.Times Books, New York.
Nicolelis M and Ribeiro S (2006) Seeking the Neural Code. Scientific American, 295, 70-77.
O’Keefe J, Burgess N (1996) Geometric determinants of the place fields of hippocampal neurons. Nature, 381, 425-428.
O’Keefe J, Burgess N (2005) Dual phase and rate coding in hippocampal place cells: theoretical significance and relationship to entorhinal grid cells. Hippocampus, 15, 853-866.
Papoutsi M, de Zwart JA, Jansma JM, Pickering MJ, Bednar JA and Horwitz B (2009) From Phonemes to Articulatory Codes: An fMRI Study of the Role of Broca’s Area in Speech Production. Cerebral Cortex,19, 2156 – 2165.
Pelc SR and Weldon MGE (1966) Stereochemical relationship between coding triplets and amino-acids. Nature, 209, 868-870.
Pertea M, Mount SM, Salzberg SL (2007) A computational survey of candidate exonic splicing enhancer motifs in the model plant Arabidopsis thaliana. BMC Bioinformatics, 8, 159.
Ray A, van der Goes van Naters W, Shiraiwa T and Carlson JR (2006) Mechanisms of Odor Receptor Gene Choice in Drosophila. Neuron, 53, 353-369.
Redies C and Takeichi M (1996) Cadherine in the developing central nervous system: an adhesive code for segmental and functional subdivisions. Developmental Biology, 180, 413-423.
Ruiz i Altaba A, Nguien V and Palma V (2003) The emergent design of the neural tube: prepattern, SHH morphogen and GLI code.Current Opinion in Genetics & Development, 13, 513–521.
Schimmel P (1987) Aminoacyl tRNA synthetases: General scheme of structure-function relationship in the polypeptides and recognition of tRNAs. Ann. Rev. Biochem., 56, 125-158.
Schimmel P, Giegé R, Moras D and Yokoyama S (1993) An operational RNA code for amino acids and possible relationship to genetic code. Proceedings of the National Academy of Sciences USA, 90, 8763-8768.
Shapiro L and Colman DR (1999) The Diversity of Cadherins and Implications for a Synaptic Adhesive Code in the CNS. Neuron, 23, 427-430.
Shimizu M (1982) Molecular basis for the genetic code. J. Mol. Evol., 18, 297-303.
Strahl BD and Allis D (2000) The language of covalent histone modifications. Nature, 403, 41-45.
Tavares EQP and Buckeridge MS (2015) Do plant cells have a code? Plant Science, 241, 286-294.
Trifonov EN (1987) Translation framing code and frame-monitoring mechanism as suggested by the analysis of mRNA and 16s rRNA nucleotide sequence. Journal of Molecular Biology, 194, 643-652.
Trifonov EN (1989) The multiple codes of nucleotide sequences. Bulletin of Mathematical Biology, 51: 417-432.
Trifonov EN (1999) Elucidating Sequence Codes: Three Codes for Evolution. Annals of the New York Academy of Sciences, 870, 330-338.
Tseng AS and Levin M (2013) Cracking the bioelectric code. Probing endogenous ionic controls of pattern formation. Communicative & Integrative Biology, 6(1), 1–8.
Turner BM (2000) Histone acetylation and an epigenetic code. BioEssays, 22, 836–845.
Turner BM (2002) Cellular memory and the Histone Code. Cell, 111, 285-291.
Verhey KJ and Gaertig J (2007) The Tubulin Code. Cell Cycle, 6 (17), 2152-2160.
Wang Z and Burge C (2008) Splicing regulation: from a part list of regulatory elements to an integrated splicing code. RNA, 14, 802-813.
Yarus M (1988) A specific amino acid binding site composed of RNA. Science, 240, 1751-1758.
Yarus M (1998) Amino acids as RNA ligands: a direct-RNA-template theory for the code’s origin. J. Mol. Evol.,47(1), 109–117.
Yarus M, Caporaso JG, and Knight R (2005) Origins of the Genetic Code: The Escaped Triplet Theory. Annual Review of Biochemistry, 74,179-198.
CODE BIOLOGY, PEIRCEAN BIOSEMIOTICS, AND ROSEN’S RELATIONAL BIOLOGY
The classical theories of the genetic code claimed that its coding rules were determined by chemistry—either by stereochemical affinities or by metabolic reactions—but the experimental evidence has revealed a totally different reality: it has shown that any codon can be associated with any amino acid, thus proving that there is no necessary link between them. The rules of the genetic code, in other words, obey the laws of physics and chemistry but are not determined by them. They are arbitrary, or conventional, rules. The result is that the genetic code is not a metaphorical entity, as implied by the classical theories, but a real code, because it is precisely the presence of arbitrary rules that divides a code from all other natural processes. In the past 20 years, furthermore, various independent discoveries have shown that many other organic codes exist in living systems, which means that the genetic code has not been an isolated case in the history of life. These experimental facts have one outstanding theoretical implication: they imply that in addition to the concept of information we must introduce in biology the concept of meaning, because we cannot have codes without meaning or meaning without codes. The problem is that at present we have two different theoretical frameworks for that purpose: one is Code Biology, where meaning is the result of coding, and the other is Peircean biosemiotics, where meaning is the result of interpretation. Recently, however, a third party has entered the scene, and it has been proposed that Robert Rosen’s relational biology can provide a bridge between Code Biology and Peircean biosemiotics.
Systems Biology: Biological Networks, Network Motifs, Switches and Oscillators
From Biological switches and clocks
The living cell receives signals from its environment and its own internal state, processes the information, and initiates appropriate responses in terms of changes in gene expression, cell movement, and cell growth or death. Like a digital computer, information processing within cells is carried out by a complex network of switches and oscillators, but instead of being fabricated from silicon transistors and quartz crystals, the cell’s computer is an evolved network of interacting genes and proteins. In the same way that computer design was made possible by a sophisticated theory of electronic circuitry, a basic understanding of cellular regulatory mechanisms will require a relevant theory of biomolecular circuitry. Although the ‘engineering mindset’ is sorely needed to make sense of the cell’s circuitry, the squishy, sloppy, massively parallel, analogue nature of biochemistry is so different from the solid-state, precise, sequential, digital nature of computers that the mathematical tools and intellectual biases of the solid-state physicist/electrical engineer are not entirely appropriate to unravelling the molecular logic of cell physiology. New modelling paradigms and software tools are evolving to meet the challenges of the new ‘systems biology’ of the living cell.
System Biology includes study of the following among other areas.
Gene co-expression networks (transcript–transcript association networks)
Metabolic networks
Signaling networks
Neuronal networks
Between-species interaction networks
Within-species interaction networks
Network Motifs:
Coherent Feedforward Loop (FFL)
Incoherent Feedforward Loop
Feedback Loop
Scaffold Motifs
Bi Fan
Multi Input Motifs (MIM)
Regulator Chains
Bi-Parallel
Single Input Module (SIM)
Dense Overlapping Regulon (DOR)
Biological Switches
Ultrasensitivity
Switches (Bistability)
Biological Oscillators
Clocks
Negative Feedback Only Oscillators
Repressilator
Pentilator
Goodwin Oscillator
Frazilator
Metabolator
Negative + Positive Feedback Oscillators
Meyer and Strayer model of Calcium Oscillations
van der Pol Oscillator
Fitzhugh-Nagumo Oscillator
Cyanobacteria Circadian Oscillator
Negative + Negative Feedback Oscillator
Negative and Positive + Negative Feedback cell cycle Oscillator
Fussenegger Oscillators
Smolen Oscillator
Amplified Negative Feedback Oscillators
Variable link Oscillators
Synthetic Biology study design of networks, switches, and oscillators.
From The dynamics and robustness of Network Motifs in transcription networks
Network Motifs
Even though biological systems are extremely complex, some of its complexity could be simplified. The study of a complex system in its entirety could prove impossible with current theories and technology. However, mathematical modelling has sought to distil the essence of complexity into concepts readily understandable by today’s science. One of such approaches has been reported by means of the study of pathways of interaction of biological networks. By concentrating on similar features that biological networks share, it has been recently discovered that at a cellular level, regulation and transcription Networks display certain patterns of connectivity at a much higher rate than expected in an equivalent randomized network. These recurring patterns of interaction, or network “Motifs”, can help us define bread classes of networks and their types of functional elements. In the same way, they can reveal the evolutionary aim by which they have been developed. Network Motifs can be interpreted as structures that have emerged as direct a reflection of the constraints under which the network has evolved. These network Motifs have been found in the biological networks of many systems, suggesting that they are the building blocks of transcription networks [4]. It has been suggested that in biological networks, these recurrent Network Motifs are responsible for carrying out key information processing tasks in the organism [5].
From Coupling oscillations and switches in genetic networks.
Switches (bistability) and oscillations (limit cycle) are omnipresent in biological networks. Synthetic genetic networks producing bistability and oscillations have been designed and constructed experimentally. However, in real biological systems, regulatory circuits are usually interconnected and the dynamics of those complex networks is often richer than the dynamics of simple modules. Here we couple the genetic Toggle switch and the Repressilator, two prototypic systems exhibiting bistability and oscillations, respectively. We study two types of coupling. In the first type, the bistable switch is under the control of the oscillator. Numerical simulation of this system allows us to determine the conditions under which a periodic switch between the two stable steady states of the Toggle switch occurs. In addition we show how birhythmicity characterized by the coexistence of two stable small-amplitude limit cycles, can easily be obtained in the system. In the second type of coupling, the oscillator is placed under the control of the Toggleswitch. Numerical simulation of this system shows that this construction could for example be exploited to generate a permanent transition from a stable steady state to self-sustained oscillations (and vice versa) after a transient external perturbation. Those results thus describe qualitative dynamical behaviors that can be generated through the coupling of two simple network modules. These results differ from the dynamical properties resulting from interlocked feedback loops systems in which a given variable is involved at the same time in both positive and negative feedbacks. Finally the models described here may be of interest in synthetic biology, as they give hints on how the coupling should be designed to get the required properties.
From Robust, Tunable Biological Oscillations from Interlinked Positive and Negative Feedback Loops
To test the generality of the idea that positive feedback enables an oscillator to have a tunable frequency and constant amplitude, we examined several other oscillator models, including five negative feedback–only models: (i) the Goodwin oscillator, a well-studied model relevant to circadian oscillations (18, 19); (ii) the Repressilator, a transcriptional triple-negative feedback loop constructed in Escherichia coli (20); (iii) the “Pentilator,” a Repressilator with five (rather than three) repressors; (iv) the Metabolator (21), a synthetic metabolic oscillator; and (v) the Frzilator, amodel of the control of gliding motions in myxobacteria (22). In four of the cases (Goodwin, Repressilator, Pentilator, and Metabolator), the amplitude/frequency curves were inverted U-shaped curves similar to that seen for the negative feedback–only cell cycle model (Figs. 1B and 3A). In the case of the Frzilator, the legs of the curve were truncated; the oscillator had a nonzero minimal amplitude (Fig. 3A). For all five of the negative feedback–only models, the oscillators functioned over only a narrow range of frequencies (Fig. 3A).
We also examined four positive-plus-negative feedback oscillators: (i) the van der Pol oscillator, inspired by studies of vacuum tubes (12); (ii) the Fitzhugh-Nagumo model of propagating action potentials (23, 24); (iii) the Meyer-Stryer model of calcium oscillations (25); and (iv) a model of circadian oscillations in the cyanobacterial KaiA/B/C system (26–28). In each case, we obtained a flat, wide amplitude/frequency curve (Fig. 3B). Thus, a tunable frequency plus constant amplitude can be obtained from many different positive-plusnegative feedback models; this feature is not peculiar to one particular topology or parameterization.
These findings rationalize why the positiveplus- negative feedback design might have been selected through evolution in cases where a tunable frequency and constant amplitude are important, such as heartbeats and cell cycles. However, it is not clear that an adjustable frequency would be advantageous for circadian oscillations, because frequency is fixed at one cycle per day. Nevertheless, the cyanobacterial circadian oscillator appears to rely on positive feedback (26), and positive feedback loops have been postulated for other circadian oscillators as well (Table 1). This raises the question of whether the positiveplus- negative feedback design might offer additional advantages.
One possibility is that the positive-plusnegative feedback design permits oscillations over a wider range of enzyme concentrations and kinetic constant values, making the oscillator easier to evolve and more robust to variations in its imperfect components. We tested this idea through a Monte Carlo approach.We formulated three simple oscillatormodels: (i) a three-variable triple negative feedback loop with no additional feedback (Fig. 4A), (ii) one with added positive feedback (Fig. 4B), or (iii) one with added negative feedback (Fig. 4C). We generated random parameter sets for the models and then for each set determined whether the model produced limit cycle oscillations.We continued generating parameter sets until we had amassed 500 that gave oscillations.
From Robust, Tunable Biological Oscillations from Interlinked Positive and Negative Feedback Loops
Key Terms:
Ultra-sensitivity
Bi-stability
Positive Feedback Loop
Negative Feedback Loop
Biological Oscillators
Biological Switches
Biological Networks
Network Motifs
Regulation Networks
Signalling Networks
Communication Networks
Biological Clocks
Circadian Rhythms
Harmonic Oscillators
Van der Pol Oscillator (Limit Cycle)
FitzHugh–Nagumo oscillators (Neural)
Limit Cycle Oscillator
Cell Cycle
Systems Biology
Synthetic Biology
Gene Regulatory Networks
Kuramoto Oscillators
Phase Coupled Oscillators
Cardic Pacemaker
Biochemical Networks
Synchronization
Goodwin Oscillator
Repressilators
Fussenegger Oscillators
Smolen Oscillators
Variable Link Oscillators
Metabolators
Amplified Negative Feedback Oscillators
Key Sources of Research:
Ultrasensitivity Part I: Michaelian responses and zero-order ultrasensitivity
Formation of Regulatory Patterns During Signal Propagation in a Mammalian Cellular Network
Avi Ma’ayan, Sherry L. Jenkins, Susana Neves, Anthony Hasseldine, Elizabeth Grace, Benjamin Dubin-Thaler, Narat J. Eungdamrong, Gehzi Weng, Prahlad T. Ram, J. Jeremy Rice, Aaron Kershenbaum, Gustavo A. Stolovitzky, Robert D. Blitzer, and Ravi Iyengar
How does human body forms from Molecules? Is it all evolutionary? or is there a role for Vitalism?
How to integrate decision making in organizations at multi levels? From Corporate level to Plant Level.
How does an Individual fits in Groups, Communities, Society, and Ecosystem?
What is the role of fractals thinking in Evolutionary Biology?
A SUMMARY OF THE PRINCIPLES OF HIERARCHY THEORY
The Hierarchy theory is a dialect of general systems theory. It has emerged as part of a movement toward a general science of complexity. Rooted in the work of economist, Herbert Simon, chemist, Ilya Prigogine, and psychologist, Jean Piaget, hierarchy theory focuses upon levels of organization and issues of scale. There is significant emphasis upon the observer in the system.
Hierarchies occur in social systems, biological structures, and in the biological taxonomies. Since scholars and laypersons use hierarchy and hierarchical concepts commonly, it would seem reasonable to have a theory of hierarchies. Hierarchy theory uses a relatively small set of principles to keep track of the complex structure and a behavior of systems with multiple levels. A set of definitions and principles follows immediately:
Hierarchy: in mathematical terms, it is a partially ordered set. In less austere terms, a hierarchy is a collection of parts with ordered asymmetric relationships inside a whole. That is to say, upper levels are above lower levels, and the relationship upwards is asymmetric with the relationships downwards.
Hierarchical levels: levels are populated by entities whose properties characterize the level in question. A given entity may belong to any number of levels, depending on the criteria used to link levels above and below. For example, an individual human being may be a member of the level i) human, ii) primate, iii) organism or iv) host of a parasite, depending on the relationship of the level in question to those above and below.
Level of organization: this type of level fits into its hierarchy by virtue of set of definitions that lock the level in question to those above and below. For example, a biological population level is an aggregate of entities from the organism level of organization, but it is only so by definition. There is no particular scale involved in the population level of organization, in that some organisms are larger than some populations, as in the case of skin parasites.
Level of observation: this type of level fits into its hierarchy by virtue of relative scaling considerations. For example, the host of a skin parasite represents the context for the population of parasites; it is a landscape, even though the host may be seen as belonging to a level of organization, organism, that is lower than the collection of parasites, a population.
The criterion for observation: when a system is observed, there are two separate considerations. One is the spatiotemporal scale at which the observations are made. The other is the criterion for observation, which defines the system in the foreground away from all the rest in the background. The criterion for observation uses the types of parts and their relationships to each other to characterize the system in the foreground. If criteria for observation are linked together in an asymmetric fashion, then the criteria lead to levels of organization. Otherwise, criteria for observation merely generate isolated classes.
The ordering of levels: there are several criteria whereby other levels reside above lower levels. These criteria often run in parallel, but sometimes only one or a few of them apply. Upper levels are above lower levels by virtue of: 1) being the context of, 2) offering constraint to, 3) behaving more slowly at a lower frequency than, 4) being populated by entities with greater integrity and higher bond strength than, and 5), containing and being made of – lower levels.
Nested and non-nested hierarchies: nested hierarchies involve levels which consist of, and contain, lower levels. Non-nested hierarchies are more general in that the requirement of containment of lower levels is relaxed. For example, an army consists of a collection of soldiers and is made up of them. Thus an army is a nested hierarchy. On the other hand, the general at the top of a military command does not consist of his soldiers and so the military command is a non-nested hierarchy with regard to the soldiers in the army. Pecking orders and a food chains are also non-nested hierarchies.
Duality in hierarchies: the dualism in hierarchies appears to come from a set of complementarities that line up with: observer-observed, process-structure, rate-dependent versus rate-independent, and part-whole. Arthur Koestler in his “Ghost in The Machine” referred to the notion of holon, which means an entity in a hierarchy that is at once a whole and at the same time a part. Thus a holon at once operates as a quasi-autonomous whole that integrates its parts, while working to integrate itself into an upper level purpose or role. The lower level answers the question “How?” and the upper level answers the question, “So what?”
Constraint versus possibilities: when one looks at a system there are two separate reasons behind what one sees. First, it is not possible to see something if the parts of the system cannot do what is required of them to achieve the arrangement in the whole. These are the limits of physical possibility. The limits of possibility come from lower levels in the hierarchy. The second entirely separate reason for what one sees is to do with what is allowed by the upper level constraints. An example here would be that mammals have five digits. There is no physical reason for mammals having five digits on their hands and feet, because it comes not from physical limits, but from the constraints of having a mammal heritage. Any number of the digits is possible within the physical limits, but in mammals only five digits are allowed by the biological constraints. Constraints come from above, while the limits as to what is possible come from below. The concept of hierarchy becomes confused unless one makes the distinction between limits from below and limits from above. The distinction between mechanisms below and purposes above turn on the issue of constraint versus possibility. Forget the distinction, and biology becomes pointlessly confused, impossibly complicated chemistry, while chemistry becomes unwieldy physics.
Complexity and self-simplification: Howard Pattee has identified that as a system becomes more elaborately hierarchical its behavior becomes simple. The reason is that, with the emergence of intermediate levels, the lowest level entities become constrained to be far from equilibrium. As a result, the lowest level entities lose degrees of freedom and are held against the upper level constraint to give constant behavior. Deep hierarchical structure indicates elaborate organization, and deep hierarchies are often considered as complex systems by virtue of hierarchical depth.
Complexity versus complicatedness: a hierarchical structure with a large number of lowest level entities, but with simple organization, offers a low flat hierarchy that is complicated rather than complex. The behavior of structurally complicated systems is behaviorally elaborate and so complicated, whereas the behavior of deep hierarchically complex systems is simple.
Hierarchy theory is as much as anything a theory of observation. It has been significantly operationalized in ecology, but has been applied relatively infrequently outside that science. There is a negative reaction to hierarchy theory in the social sciences, by virtue of implications of rigid autocratic systems or authority. When applied in a more general fashion, even liberal and non-authoritarian systems can be described effectively in hierarchical terms. There is a politically correct set of labels that avoid the word hierarchy, but they unnecessarily introduce jargon into a field that has enough special vocabulary as it is.
A SHORT ANNOTATED BIBLIOGRAPHY OF HIERARCHY THEORY.
This bibliography is in chronological order, so that the reader can identify the early classics as opposed to the later refinements. If you must choose just one book to read, turn to the last reference in this bibliography, Ahl and Allen, 1996. Simon, H.. A. 1962. The architecture of complexity. Proceedings of the American philosophical society 106: 467-82. This is the foundation paper of hierarchy theory originating from an economist. It was a re-published in “Sciences of the Artificial” by Simon. It introduces the idea of near-decomposability. If systems were completely decomposable, then there would be no emergent whole, because the parts would exist only separately. The “near” in near-decomposable allows the upper level to emerge from the fact that the parts anre not completely separate.
Koestler, Arthur. 1967. The ghost in the machine. Macmillan, New York. This is a long hard look at human social structure in hierarchical terms. The notion of holon first occurs in this work. This is a classic work, but is easily accessible to the lay public.
Whyte, L.. L.., A. G. Wilson and D. Wilson (eds.). 1969. Hierarchical structures. American Elsevier, New York. This is a classic collection of early scholarly works by some of the founders of hierarchical thinking.
Pattee, H.. H. (ed.) 1973. Hierarchy theory: the challenge or complex systems. Braziller, New York. This edited volume has some classic articles by Pattee, Simon and others.
Allen, T. F. H. and T. B. Starr. 1982. Hierarchy: perspectives for ecological complexity. University Chicago Press. This book has a significant ecological component but is much more generally about hierarchical structure. It is abstract and a somewhat technical treatment but has been the foundation work for the application of hierarchy theory in ecology and complex systems theory at large.
Salthe, S. 1985. Evolving Hierarchical Systems: their structure and representation. Columbia University Press, New York. This book has a strong structural bias, in contrast to the process oriented approach of Allen and the other ecologists in this bibliography. Salthe introduces the notion of the Triadic, where there is a focus on 1) the system as both a whole above the levels below and 2) a part belonging to another level above, 3) not forgetting the level of the structure itself in between. While much biological hierarchy theory takes an anti-realist point view, or is at least reality-agnostic, wherein the ultimate reality of hierarchical arrangement is left moot, Salthe’s version of hierarchy theory is concerned with the ultimate reality of structure. The anti-realist view of structure is that it is imposed by the observer, and may or may not correspond to any ultimate reality. If structure does correspond to ultimate, external reality, we could never know that to be so. Salthe’s logic is consistent but always takes a structural and ontological position.
O’Neill, R. V., D. DeAngelis, J. Waide and T. F. H. Allen. 1986. A hierarchical concept of ecosystems. Princeton University Press. This is a distinctly ecological application of hierarchy theory, making the critical distinction between process functional ecosystem approaches as opposed to population and community relationships. It is an application of hierarchy theory to ecosystem analysis.
Allen T. F. H. and T. Hoekstra. 1992. Toward a unified ecology. Columbia University Press. This book turns on hierarchy theory, but is principally a book about ecology. It goes beyond the O’Neill et al book, in that it makes the distinction between many types of ecology (landscape, ecosystem, community, organism, population, and biomes) on the one hand, and scale of ecology on the other hand. It ends with practical applications of hierarchy theory and ecological management.
Ahl, V. and T. F. H. Allen. 1996. Hierarchy theory, a vision, vocabulary and epistemology. Columbia University Press. This slim a volume is an interdisciplinary account of a hierarchy theory, and represents the shallow end of the pool. It is the primer version of Allen and Starr 1982. It is full of graphical images to ease the reader into a hierarchical perspective. It makes the distinction between levels of organization and levels of observation. It takes a moderate anti-realist point of view, wherein there may be an external reality, but it is not relevant to the discourse. We only have access to experience, which must of necessity involve observer values and subjectivity. There are examples from a wide discussion of many disciplines. Included are examples from psychology, ecology, the law, political systems and philosophy. It makes reference to the global and technological problems facing humanity, and offers hierarchy theory as one tool in the struggle. The summary of hierarchy theory in the opening paragraphs above comes from this book.
This summary was compiled by
Timothy F. Allen, Professor of Botany,
University of Wisconsin Madison,
Madison Wisconsin 53706 — 1381.
Email – tfallen@facstaff.wisc.edu
On Anticipation: Going Beyond Forecasts and Scenarios
From Anticipation.Info of Mihai Nadin
A Second Cartesian Revolution
For about 400 years, humankind, or at least the western world, has let itself be guided by the foundation set by Descartes and Newton. The cause-and-effect, deterministic model of the machine became so powerful that every thing and every being came to be considered a machine. As a description of the material world and as an expression of the laws governing its functioning, deterministic-based physics and Cartesian reductionism (of the whole to its parts) proved to be extremely powerful instruments in the overall progress of humankind. But neither Descartes nor Newton, nor most of their followers, could have envisioned the spectacular development of science in its current depth and breadth.
The physicist Erwin Schrödinger concluded that organisms are subject to “a new physics,” which he did not produce, but rather viewed as necessary. This new physics might well be the domain of anticipation. Indeed, from within physics itself—that is, quantum mechanics—a possible understanding of some aspects of anticipation can be derived.
The realization that the world is the unity of reaction and anticipation is not new. What is new is the awareness of the limits of our understanding a dynamics of change that transcends the deterministic view. The urgent need for such an understanding is probably best expressed in the spectacular development of the life sciences.
The perspective of the world that anticipation opens justifies the descriptor “a second Cartesian Revolution.” Instead of explaining complexity away, we will have to integrate it into our existence as the informational substratum of rich forms through which anticipatory processes take place.
From Anticipation.Info of Mihai Nadin
Anticipation: Why is it a subject of research?
Anticipation occurs in all spheres of life. It complements the physics of reaction with the pro-active quality of the living. Nature evolves in a continuous anticipatory fashion targeted at survival. The dynamics of stem cells demonstrate this mechanism. Through entailment from a basic stem cell an infinite variety of biological expression becomes possible.
Sometimes we humans are aware of anticipation, as when we plan. Often, we are not aware of it, as when processesembedded in our body and mind take place before we realize their finality. In tennis, for example, the return of a professional serve can be successful only through anticipatory mechanisms. A conscious reaction takes too long to process. Anticipation is the engine driving the stock market. Creativity in art and design are fired by anticipation.
“The end is where we start from,” T. S. Eliot once wrote. Before the archer draws his bow, his mind has already hit the target. Motivation mechanisms in learning, the arts, and all types of research are dominated by the underlying principle that a future state—the result—controls present action, aimed at success. The entire subject of prevention entails anticipatory mechanisms.
From Anticipation.Info of Mihai Nadin
Research into anticipation revealed various aspects that suggested a number of definitions.
Robert Rosen, Mihai Nadin, Daniel Dennett and others who approached particular aspects of anticipation contributed to some of these definitions. Mihai Nadin (cf. Anticipation – A Spooky Computation) attempted an overview of the various angles from which anticipation can be approached if the focus is on computation. This overview is continued and expanded in the integrated publication (book+dvd+website) to which this website belongs. The following 12 definitions, or descriptions, of anticipation should be understood as working hypotheses. It is hoped and expected that the knowledge community of those interested in anticipation will eventually refine these definitions and suggest new ones in order to facilitate a better understanding of what anticipation is and its importance for the survival of living systems.
An anticipatory system is a system whose current state is determined by a future state. “The cause lies in the future,”. (cf. Robert Rosen, Heinz von Foerster)
Anticipation is the generation of a multitude of dynamic models of human actions and the resolution of their conflict. (cf. Mihai Nadin)
An anticipatory system is a system containing a predictive model of itself and/or of its environment that allows it to change state at an instant in accord with the model’s predictions pertaining to a later instant. (cf. Robert Rosen)
Anticipation is a process of co-relation among factors pertaining to the present, past and future of a system. (cf. Mihai Nadin)
Anticipation is an expression of the connectedness of the world, in particular of quantum non-locality. (cf. Mihai Nadin)
Anticipation is the expression of natural entailment. (cf. Robert Rosen)
Anticipation is a mechanism of synchronization and integration. (cf. Mihai Nadin)
Anticipation is an attractor within dynamic systems. (cf. Mihai Nadin)
Anticipation is a recursive process described through the functioning of a mechanism whose past, present, and future states allow it to evolve from an initial to a final state that is implicitly embedded in the mechanism. (cf. Mihai Nadin)
Anticipation is a realization within the domain of possibilities. (cf. Mihai Nadin)
Anticipatory mechanisms can be reinforced through feedback. Feedforward and inverse kinetics are part of the integrated mechanism of anticipation. (cf. Daniel Dennett, Daniel Wolpert, Nadin)
Anticipation is a power law-based long-range interaction. (cf. Mihai Nadin)
From An Introduction to the Ontology of Anticipation
Recent years have witnessed the growth of significant interest in theories and methodologies which seek to foresee the future development of relevant situations. Studies of the future fall under many different denominations, and they employ a huge variety of techniques, ranging from forecasting to simulation, from planning to trend extrapolation, from future studies and scenarios to anticipatory systems. Widely different conceptualisations and formalisations have been proposed as well.1 This remarkable variety may be partly simplified by making explicit the main underlying assumptions of at least some of them. Two of these assumptions are that (1) the future is at least partly governed by the past, and (2) the future can be better confronted by opening our minds and learning to consider different viewpoints. According to (1) the future is part of a structured story whose past and present are at least partially known. The claim is defended that the forces that have shaped past and present situations will still be valid while the situation under consideration unfolds. The core thesis is that the future is embedded in the past; it is the projection of the past through the present. Time series analysis, trend extrapolation, and forecasting pertain to this family. Any of the mentioned methodologies may be further supplemented by computer-based simulations. On the other hand, instead of directly addressing the problem of searching for the seeds of the future in the past, (2) considers the different problem of preparing for the unforeseeable novelties awaiting us in the future. Learning about widely different outcomes is now the issue: one must be ready to consider and address possibly unfamiliar or alien scenarios. The main outcome of this exercise is an increased capacity to distinguish among possible, probable, and preferred future scenarios. These activities come under the heading of future studies, while scenario construction is the best known methodology adopted by practitioners. For now on I shall refer to (1) and (2) as respectively the forecasting and the scenario viewpoints. Forecasts and scenarios are not contradictory one to the other. They may and usually do coexist, since they address the future from two different standpoints. Furthermore, experience shows that both are useful. This paper introduces a third, different viewpoint, here termed the viewpoint of anticipatory systems, which can be profitably synthesized with forecasts and scenarios; i.e. it is not contradictory with the claims of either the forecasting or scenario viewpoint. Recent years have witnessed the growth of significant interest in anticipation.2 Anticipatory theories have been proposed in fields as different as physics, biology, physiology, neurobiology, psychology, sociology, economy, political science, computer science and philosophy. Unfortunately, no systematic comparison among the different viewpoints has so far been developed. It is therefore fair to claim that currently no general theory of anticipation is available. Generally speaking, anticipation concerns the capacity exhibited by some systems to tune their behaviour according to a model of the future evolution of the environment in which they are embedded. Generally speaking, the thesis is defended that “An anticipatory system is a system containing a predictive model of itself and/or its enviroment, which allows it to change state at an instant in accord with the model‟s predictions pertaining to a later instant” (Rosen [19: 341]). The main difference between forecasting and scenarios on the one hand, and anticipation on the 1 See, among many others, Adam [1], Bell [4], Cornish [5], Godet [7], Lindgren and Bandhold [8], Retzbach [16], Slaughter [22], Woodgate and Pethrick [23]. 2 Starting from the seminal Rosen [19]. See also [20], [21]. 2 other, is that the latter is a property of the system, intrinsic to its functioning, while the former are cognitive strategies that a system A develops in order to understand the future of some other system B (of which A may or may not be a component element).
Key Terms
Hyper Sets
Hyper Incursion
Hyper Recursion
Recursion
Incursion
Anticipatory Systems
Weak Anticipation
Strong Anticipation
Key People
Roberto Poli
Mihai Nadin
Riel Miller
Robert Rosen
John J Kineman
Daniel M Dubois
John Collier
Loet Leydesdorff
Key Sources of Research:
Systems and models with anticipation in physics and its applications

Figure 7 When I made this photograph, the structure was enclosed in a mirrored pyramid.
Figure 8 One sphere is removed from the outer cluster to reveal the inner cluster. However, the inner cluster must be upside down with respect to the outer cluster to fit inside.
Figure 36 Zoom-in This is the image on the forehead of the face in Figure 36 Large.
Figure 36 Very wide-angle Large



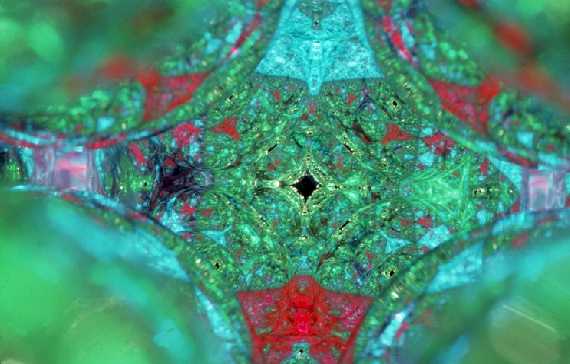
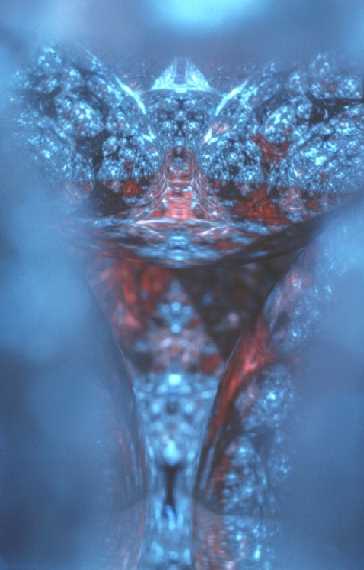


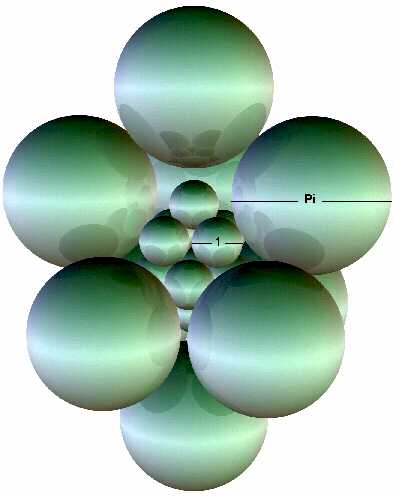



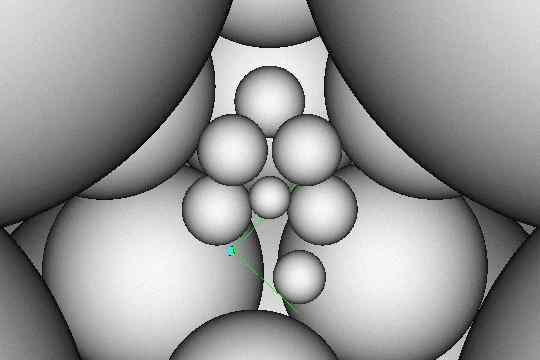









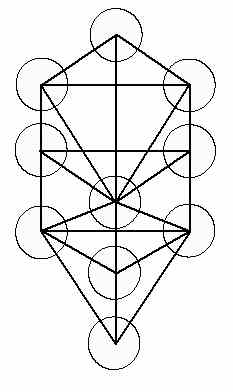



















.jpg)
























 , 1, … crossings are 1, 0, 0, 1, 1, 2, 5, 8, 25, … (OEIS
, 1, … crossings are 1, 0, 0, 1, 1, 2, 5, 8, 25, … (OEIS  . It has since been shown that a knot cannot exist in any dimension
. It has since been shown that a knot cannot exist in any dimension  . Two distinct knots cannot have the same
. Two distinct knots cannot have the same  crossings is a member of the
crossings is a member of the  .
.





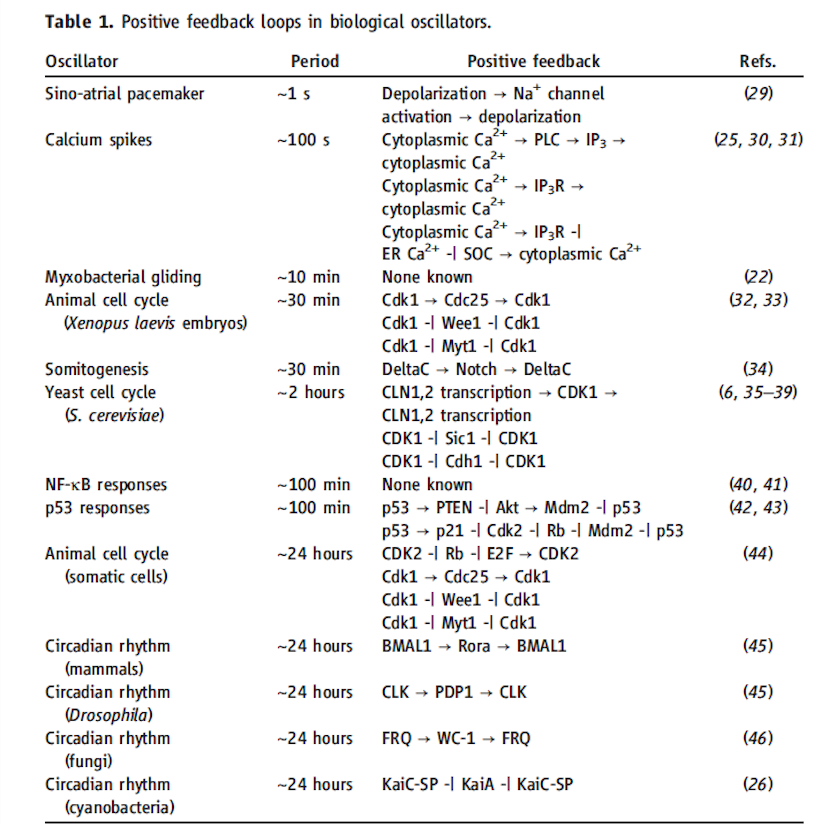

{kind=link}
{kind=link}
{kind=link}
{kind=link}